
凹叶厚朴种子休眠与萌发特性的研究
2011-12-23 03:07:54胡江琴冯晓恩沈檬笑
杭州师范大学学报(自然科学版) 2011年4期
胡江琴,冯晓恩,沈檬笑,王 赟
(杭州师范大学生命与环境科学学院,浙江 杭州 310036)
凹叶厚朴种子休眠与萌发特性的研究
胡江琴,冯晓恩,沈檬笑,王 赟
(杭州师范大学生命与环境科学学院,浙江 杭州 310036)
用组织切片法对凹叶厚朴种子胚形态结构的观察、种皮透水性的测定、种子不同部位浸提液的生物测定,初步探讨了凹叶厚朴种子的休眠原因.结果表明:凹叶厚朴种子的胚已分化完全;内种皮有阻碍种子吸水的作用;外种皮和中种皮以及胚乳存在较多的发芽抑制物质.对凹叶厚朴种子的萌发试验表明,用1 500mg/L赤霉素(GA3)浸种后低温层积处理是解除休眠、促进萌发的最佳方法,种子发芽率可达72%,比单独低温层积处理提高了17%.
凹叶厚朴;种子;休眠;萌发
种子休眠是植物本身适应环境和延续生存的一种特性,是种子植物进化的一种稳定对策.野生植物特别是原产温带的植物,其种子大多有深而长的休眠期.凹叶厚朴(Magnolia biloba)是木兰科木兰属落叶乔木,原产四川、湖北、湖南、广西、浙江、江西、福建、江苏等地[1].其集药用、材用、观赏于一体,是我国特有的经济树种,也是国家二级重点保护植物[2],利用前景十分广阔.多年来由于凹叶厚朴需求量不断增加,野生资源不断减少,而凹叶厚朴种子又具有后熟性和硬实性[1],具有深休眠特性,自然繁殖率低.研究和探讨凹叶厚朴种子的休眠和萌发,将为扩大凹叶厚朴资源、更好地保护和利用这一优良树种提供科学的依据.该文对凹叶厚朴种子休眠与萌发的特性进行研究,以期找到其休眠的主要原因及打破休眠的有效方法.
1 材料与方法
1.1 材 料
该试验中种子不同部位浸提液生物测定所用的凹叶厚朴种子(带红色外种皮),于2008年11月采自杭州植物园.其他试验所用的凹叶厚朴种子(除去外种皮和中种皮),于2008年11月采购自广西龙胜县伟江乡.白菜种子为“早熟五号”一代交配白菜种子,采购自杭州种子公司.
1.2 方 法
1.2.1 胚形态结构观察
选取颗粒饱满的种子,剥去种皮,用FAA固定,常规石蜡切片,厚度为8μm,番红-固绿染色,在Zeiss荧光显微镜下观察胚的形态结构并拍照.
1.2.2 种皮透水性测定
取完整种子(即已脱去红色革质假种皮和肉质中种皮,保留内种皮的凹叶厚朴种子,下面统称为完整种子)、破皮种子(即剥去内种皮,保留胚和胚乳的凹叶厚朴种子,下面统称为破皮种子)各20粒,分别称其干重,放入小烧杯中,加入25℃温水50mL浸种,搅拌至冷却.分别于2、4、24、48、64、72、88、92、108h取出种子,用吸水纸吸干种子表面水分,用1/10 000电子天平称重并记录.然后再把种子放回小烧杯,加50mL蒸馏水浸泡,称重.直至2次测定的值处于接近(恒重)为止,计算吸水率,每个处理设3个重复[3].

1.2.3 种子不同部位浸提液的生物测定
取种子的不同部位(外种皮+中种皮、内种皮、胚乳)各2.7g,加80%甲醇研磨,4℃密封浸提48h,其间振荡5次,过滤,将滤液在35℃下减压浓缩蒸干,并定容至40mL.然后将浸提液分别稀释为原液的20%、50%和100%.在直径为9cm的培养皿中放置滤纸,分别加入上述不同浓度的浸提液3mL,以同体积蒸馏水为对照.每种处理设4个重复,每个重复放置50粒白菜种子,于25℃恒温光照培养箱内培养,24h测定白菜种子的发芽率(以露出子叶为发芽的标准),48h测量胚根长度[3-4].
1.2.4 层积处理
层积基质为含水量约60%的湿沙(即手握成团、松手即散[5]),基质在使用前130℃高温杀菌1h.选取颗粒饱满种子,将其与湿沙混匀(湿沙与种子比例为3∶1),然后分别在低温(4℃)、室温、变温(13℃/16h、4℃/8h)3种条件下进行层积.每种处理设3个重复.
1.2.5 赤霉素(GA3)浸种后低温层积处理
选取颗粒饱满种子,先分别用浓度为500、1 000、1 500mg/L的赤霉素(GA3)溶液浸种36h,然后进行低温(4℃)层积.每种处理设3个重复.
1.2.6 发芽试验
将上述经过处理的种子,于层积90d后,用30℃温水浸泡24h,0.5%高锰酸钾灭菌10min,置于种子萌发盒中在25℃温度下萌发,萌发第12天测定种子发芽率(以露白为准),测量胚根长度.
1.2.7 数据处理
利用Microsoft Excel电子表格软件对所得实验数据进行方差分析,采用Duncan's新复极差法进行多重比较,并用字母标记法标记其差异性.
2 结果与分析
凹叶厚朴种子包括种皮、胚乳、胚3个部分;种皮由外种皮、中种皮和内种皮组成;外种皮革质,中种皮肉质,内种皮木质;胚小,胚乳丰富[6].
2.1 胚的形态结构
凹叶厚朴果实脱落时种子的胚已分化完全,具有明显的子叶、胚芽、胚轴及胚根,胚的形态已处于鱼雷形阶段(见图1).由此可以判断,凹叶厚朴种子休眠的原因,不是因为胚未分化完全,而是生理上未成熟,其休眠类型为生理休眠.这一结果与天女木兰种子休眠的主要原因(胚未发育完全[7])不同.
2.2 种皮透水性对种子休眠的影响
通过定时测定完整种子和破皮种子的水分变化情况,绘出种子吸水过程的变化曲线(见图2).

图1 凹叶厚朴种子的显微结构(示胚的形态)200×Fig.1 Microstructure of the seeds of Magnolia biloba(showing embryo)200×
从图2可以看出,破皮种子的吸水特点:前48h吸水率直线上升,吸水速度较快,达到12.7%;48~88h,吸水速度开始减慢,直至88h吸水达到饱和,其吸水率达到最大值为14.2%;88h以后吸水率有所降低,108h吸水率为14.0%,吸水率在达到最大值后略有降低可能是由于长时间在水中浸泡导致种子内含物外渗的结果[3].
完整种子的吸水特点:前48h吸水率直线上升,吸水速度较快,达到8.6%;48~72h,吸水速度开始减慢,直至72h吸水达到饱和,其吸水率达到最大值为9.4%;72h以后,吸水率基本保持不变.
对比发现,完整种子与破皮种子的吸水率有较大差异,破皮种子的吸水率明显大于完整种子.这说明坚硬的内种皮有阻碍种子吸水的作用.而破皮种子打破了内种皮的吸水阻碍作用,从而明显提高了其吸水率.这也说明了坚硬的内种皮可能是导致种子休眠的原因之一.
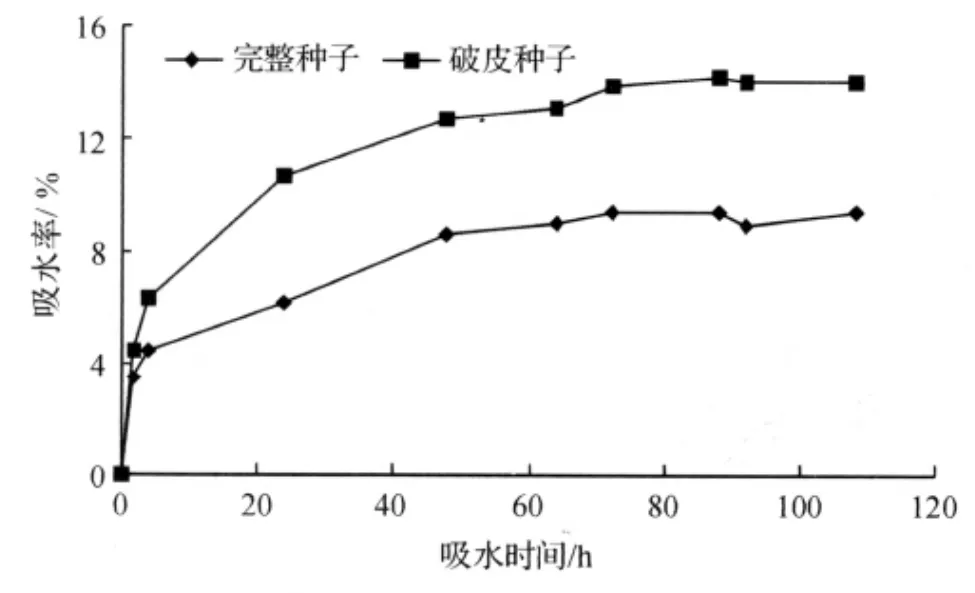
图2 凹叶厚朴种子的吸水曲线Fig.2 Water absorption curve of the seeds of Magnolia biloba
在导致种子生理休眠的诸多原因中,抑制物质被认为是最重要的原因之一,发芽抑制物质在植物种子中广泛存在[8].凹叶厚朴种子外种皮+中种皮、内种皮、胚乳浸提液对白菜种子发芽均有一定的抑制作用(见表1).方差分析结果表明,不同部位不同浓度的浸提液与对照的白菜种子发芽率的差异均达到了显著水平.外种皮+中种皮和胚乳浸提液对白菜种子的抑制作用比内种皮强得多,外种皮+中种皮抑制作用最强,并且随着浸提液浓度的降低,白菜种子的发芽率逐渐升高,说明抑制作用逐渐减弱.

表1 凹叶厚朴种子不同部位浸提液对白菜种子发芽和生长的影响Tab.1 Effects of extract in different parts from the seeds of Magnolia biloba on germination and growth of cabbage seeds
2.3 种子不同部位浸提液对白菜种子发芽和生长的影响
方差分析结果表明,凹叶厚朴种子不同部位不同浓度浸提液处理白菜种子,萌发得到的胚根长度与对照相比有显著差异,说明浸提液对白菜种子胚根的生长均有一定的抑制作用,外种皮+中种皮的抑制作用最强,而且浸提液浓度越高这种抑制作用越强.
从分析结果可以推断,凹叶厚朴种子外种皮+中种皮、内种皮、胚乳都含有发芽抑制物质,对白菜种子的发芽和生长均有一定抑制作用,且抑制程度因抑制部位而有所不同.这可能是造成凹叶厚朴种子休眠的另一个原因.
2.4 层积处理对凹叶厚朴种子发芽的影响
层积处理可以使种皮软化,增加透性,消除发芽抑制物质,是人们常用的解除休眠的有效方法.李铁华等[9]对楠木种子休眠的研究表明,层积处理能逐步解除楠木种子的休眠.层积的温度因种而异.王利宝等[10]认为对解除油松种胚休眠效果来说,低温层积处理是最好的方法.凹叶厚朴种子与湿沙在不同温度下层积,结果见表2.低温(4℃)层积、变温(13℃/4℃)层积、室温层积处理后,发现低温层积处理后的种子发芽率最高.经多重比较得出,低温层积、变温层积处理的发芽率和室温层积处理差异均达显著水平,而低温层积与变温层积处理的发芽率差异不显著.

表2 层积处理对凹叶厚朴种子发芽的影响Tab.2 Effects of stratification treatment on the seed germination of Magnolia biloba
2.5 赤霉素(GA3)浸种后低温层积对凹叶厚朴种子发芽的影响
赤霉素(GA3)是一种植物激素,通常情况下,它对许多具有休眠特性的种子的萌发有促进作用.如赖家业等[11]对单性木兰种子的研究表明,赤霉素能够打破其休眠.表3显示的是不同浓度的GA3浸种后低温层积对凹叶厚朴种子发芽的影响.经多重比较得出,不同浓度GA3浸种,凹叶厚朴种子的发芽率差异不显著.从表3可以看出,以1500mg/L GA3浸种后低温层积最为有效,种子的发芽率比单独低温层积处理(55%)提高了17%.

表3 赤霉素和低温层积对凹叶厚朴种子发芽的影响Tab.3 Effects of gibberellin and cold stratification on the seed germination of Magnolia biloba
3 结 论
通过对凹叶厚朴种子休眠特性的研究,证明胚已分化完全,其休眠类型为生理休眠;内种皮有阻碍种子吸水的作用;外种皮+中种皮、内种皮、胚乳均存在发芽抑制物质,抑制物质在外种皮+中种皮>胚乳>内种皮,且随着浸提液浓度的升高,对白菜种子的发芽率和胚根长的影响越来越明显.
层积处理能逐步解除凹叶厚朴种子的休眠,低温(4℃)层积、变温(13℃/4℃)层积比室温层积的效果好,低温层积后种子的发芽率为55%.赤霉素(GA3)浸种后低温层积,能促进发芽,其中以1500mg/L GA3浸种后低温层积最为有效,种子发芽率达72%,比单独低温层积处理提高了17%.
[1]张中玉,张建华.凹叶厚朴育苗技术[J].甘肃科技.2007,23(2):207-208.
[2]张国珍,李策宏,谢孔平,等.凹叶厚朴种子保存方法及幼苗生长规律[J].林业科技开发,2008,22(5):19-21.
[3]付晓云,于光艳,周广柱.日本厚朴种子休眠原因初探[J].辽宁林业科技,2008(1):1-3.
[4]张艳杰,高捍东,鲁顺保.南方红豆杉种子中发芽抑制物的研究[J].南京林业大学学报,2007,31(4):51-56.
[5]郭永清,沈永宝,喻方圆,等.北美鹅掌楸种子破眠技术研究[J].浙江林业科技,2006,26(6):38-40.
[6]胡江琴.凹叶厚朴种子的结构和细胞化学研究[J].杭州师范大学学报:自然科学版,2009,8(6):449-452.
[7]杜凤国,王欢,杨德冒,等.天女木兰种子形态及生物学特性[J].北华大学学报,2006,7(3):269-272.
[8]Bewley J D,Black M.Seeds:physiology of development and germination[M].New York:Plenum Press,1994:199-257.
[9]李铁华,彭险峰,喻勋林,等.楠木种子休眠与萌发特性的研究[J].中国种业,2008(1):43-45.
[10]王利宝,董丽芬.油松种胚休眠特性及解除胚休眠的方法[J].中南林学院学报,2006,26(3):19-23.
[11]赖家业,刘敬宝,潘春柳,等.不同处理对单性木兰种子萌发的影响[J].广西科学,2008,15(2):195-197.
Study on the Characteristics of Seed Dormancy and Germination of Magnolia biloba
HU Jiang-qin,FENG Xiao-en,SHEN Meng-xiao,WANG Yun
(College of Life and Environmental Sciences,Hangzhou Normal University,Hangzhou 310036,China)
Using the technique of tissue slicing,the experiment observed the morphology structure of embryo,measured the permeability of seed coat and the bioassay of the extract from different parts of Magnolia bilobaseeds,and discussed the dormancy cause of Magnolia bilobaseeds.The results show that the embryo is fully differentiated,the water absorptivity of seed is impeded by endopleura,and there exist more inhibitors in the endosperm,episperm and mesosperm of seeds.The results of germination experiment on Magnolia biloba seeds show that treating the seeds of Magnolia biloba with cold stratification after immersing into 1 500mg/L GA3is the optimal method to break dormancy and promote germination,which can obtain 72%germination,and increase 17%compared to the seeds treated only with cold stratification.
Magnolia biloba;seed;dormancy;germination
Q945.53
A
1674-232X(2011)04-0329-04
10.3969/j.issn.1674-232X.2011.04.009
2011-04-13
浙江省自然科学基金项目(Y3100241).
胡江琴(1967—),女,浙江萧山人,副教授,硕士,主要从事结构植物学研究.E-mail:jq_hu1967@163.com
猜你喜欢
植物资源与环境学报(2022年6期)2022-12-16 11:35:12
防护林科技(2020年11期)2020-12-30 03:55:36
潍坊学院学报(2020年6期)2020-11-22 08:04:08
中国农业大学学报(2020年8期)2020-07-22 07:57:28
中国食品学报(2019年3期)2019-01-13 01:47:39
许昌学院学报(2018年8期)2018-09-05 02:05:18
小学阅读指南·高年级版(2016年9期)2016-10-31 05:23:01
蔬菜(2016年10期)2016-03-27 12:35:11
武夷学院学报(2015年3期)2015-07-18 11:03:47
云南中医学院学报(2014年3期)2014-07-31 18:58:03
