
万木林青年和老龄常绿阔叶林乔木层碳贮量分配特征*
2011-12-15 09:01杨智杰陈光水
亚热带资源与环境学报 2011年2期
黄 蓉,王 超,杨智杰,陈光水
(1.福建师范大学地理研究所,福州 350007;2.湿润亚热带山地生态国家重点实验室培育基地,福州 350007)
万木林青年和老龄常绿阔叶林乔木层碳贮量分配特征*
黄 蓉,王 超,杨智杰,陈光水
(1.福建师范大学地理研究所,福州 350007;2.湿润亚热带山地生态国家重点实验室培育基地,福州 350007)
了解森林生物量碳分配特征,对于森林碳贮量和碳汇估算、森林碳汇经营和生物多样性保护有重要意义.本课题选择福建省万木林自然保护区青年 (约47 a生)和老龄 (约120 a生)常绿阔叶林为研究对象,根据生物多样性调查结果和生物量异速方程,计算乔木层生物量碳贮量,并分析其高度级、径级和树种间的分配特征.结果表明:1)青年林碳贮量为115.03 t·hm-2,老龄林碳贮量为224.43 t·hm-2;2)青年林最大碳贮量出现在高度级Ⅶ级(15~17 m),为23.17 t·hm-2,而老年林最大碳贮量出现在高度级Ⅷ级 (17~19 m),为62.96 t·hm-2;3)青年林中Ⅵ级 (15 cm≤DBH<17 cm)的碳贮量最高,为34.25 t·hm-2,占总碳贮量的29.77%;在老龄林中最高碳贮量集中在XIX级 (DBH≥41 cm),为60.03 t·hm-2,占总碳贮量的26.78%;4)2个林分绝大部分碳贮量都集中在少数树种,大多数物种对碳贮量的贡献很微小.
乔木碳贮量;碳分配;常绿阔叶林;多度;丰富度
森林生物量碳库是陆地生态系统最大的生物量碳库,森林植被碳贮量约占陆地植被碳贮量的82.5%[1],是森林固碳能力的重要标志,也是森林碳收支评估的重要参数之一[2].在森林植被碳贮量中,乔木层碳贮量占大部分,是森林碳汇功能的主体.有关森林乔木层碳贮量的分配信息,包括各器官的分配 (如地上、地下部分)、空间分配 (径级、高度分配)及树种间分配等,对于深入了解森林结构与碳汇功能关系具有重要作用.林龄对森林碳贮量特别是生物量碳贮量具有显著的影响,但目前有关林龄对森林生物量碳贮量分配的影响主要集中在各器官间的分配,而对林龄如何影响森林特别是中亚热带常绿阔叶林生物量碳贮量的空间分配特征则知之甚少.有关生物多样性与碳吸存关系是国际研究热点,但相关研究主要集中在草地、农田生态系统[3-4],有关森林植物多样性与碳贮量关系的研究极少,特别是森林树种多样性 (如丰富度、多度等)对碳贮量的作用目前尚不清楚.因此,本课题选择福建万木林自然保护区青年常绿阔叶林 (天然常绿阔叶林经破坏后次生演替形成,年龄约为47 a左右)和老龄常绿阔叶林 (次生演替形成,年龄为120 a左右)为研究对象,研究青年林和老龄林乔木层碳贮量及其分配 (径级、高度和树种间分配)的差异,以期为中亚热带常绿阔叶林森林碳贮量和碳汇的科学估算、常绿阔叶次生林的碳汇经营以及生物多样性保护等提供参考.
1 研究地概况
万木林自然保护区位于福建省北部的建瓯市,地理位置为27°03'N,118°09'E,全区南北长1.82 km,东西宽1.04 km,总面积为1.89 km2.本地区属于中亚热带季风气候,多年平均气温19.4℃,多年平均降水量1 731.4 mm,多年平均蒸发量1 466 mm,相对湿度81%,全年无霜期277 d.自然保护区属于低山丘陵地带,平均海拔360 m.所选择的青年林和老龄林的林分基本情况见表1.
2 研究方法
2.1 数据来源
2009年8月,分别在青年林和老龄林中布设1 200 m2生物多样性调查样方.按乔木、灌木、草本3层进行多样性调查,其中乔木层调查树种、树高、胸径.本研究选取了DBH≥5 cm的树种估算乔木层碳贮量.
2.2 数据处理
由于树高H和胸径D等测树因子与植株生物量有密切关系,因此本研究采用“相对生长法”来估算植株的生物量,其中计算用公式W=0.183 4(D2H)0.8156(通过收集已发表的文献中常绿阔叶林林分生物量平均单株生物量及平均胸径、树高数据拟合而到公式,暂未发表),计算常绿阔叶树种的单株生物量,再乘以林分密度得到乔木层生物量.乔木层碳贮量的计算则是根据公式:乔木层碳贮量Ts=乔木层生物量W×植株含碳率%[5].其中常绿阔叶树的平均含碳率为55.49%[6].用Excel对数据进行整理和分析.
3 结果与分析
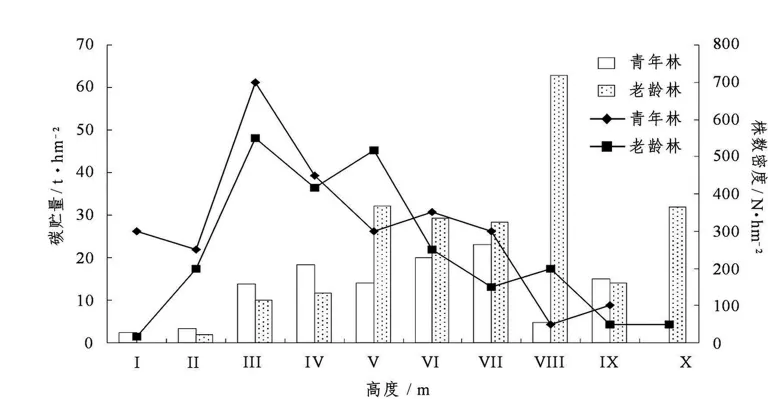
图1 不同林龄常绿阔叶林各高度级碳贮量与株数Figure 1 Height allocation of carbon storage and plant number in evergreen broad-leaved forests at different age
3.1 乔木层碳贮量的垂直分配
青年林和老龄林各高度级内的植株密度和碳贮量关系见图1.2种林分的株数随高度级增大均呈先增加后降低的趋势,总体上中低高度级的植株个体数量均占绝对优势.
2种林分的碳贮量变化趋势基本一致(图1).青年林径级Ⅰ~Ⅳ级、Ⅳ ~Ⅶ级内形成碳贮量递增的分布规律;而老年林呈现碳贮量递增规律的径级范围为Ⅰ~Ⅴ级.青年林中各高度级内树木株数呈现单峰型,高度Ⅲ级中植株密度最大,但最大碳贮量出现在Ⅶ级(15~17 m),为23.17 t·hm-2,占总碳贮量的20.14%,呈现出植株密度与和碳贮量分布不相符的情况.老龄林中各高度级内树木株数为双峰型,株数最大的高度级为Ⅲ级,株数密度为550株·hm-2,其碳贮量为10.12 t·hm-2;但老年林最大碳贮量出现在Ⅷ级 (17~19 m),植株数仅为Ⅲ级的一半,其碳贮量为62.96 t·hm-2,占总碳贮量的28.30%.
3.2 乔木层碳贮量的径级分配
图2表明2个林分株数的径级分布和碳贮量的径级分布不一致.小径级中的植株个数占主要优势.青年林5 cm≤DBH<15 cm径级内,随着胸径增加,个体数量减少,但碳贮量呈增加的趋势,这说明胸径对碳贮量的影响大于植株个数,Ⅰ级和Ⅱ级径级内个数与Ⅵ级相近,但是绝对碳贮量却远远小于Ⅵ级.老龄林中两者的分配也不一致,老龄林17 cm≤DBH<25 cm(Ⅷ级、Ⅸ级、Ⅹ级)碳贮量在DBH和株数双重作用下增加,而Ⅹ级、Ⅺ级中碳贮量随着径级和个数的减少反而增加,这显示出DBH和株数之间的相互作用.Ⅱ级、XIX级分别为植株个数的最高点和碳贮量最高点,但却不是碳贮量的最高点和植株个数的最高点,显示出DBH和株数之间的抵消作用.
在青年林中Ⅵ级 (15 cm≤DBH<17 cm)的碳贮量最高,为34.25 t·hm-2,占总碳贮量的29.77%;在老龄林中最高碳贮量集中在 XIX(DBH≥41 cm),为60.03 t·hm-2,占总碳贮量的26.78%.老龄林的碳贮量集中在一些大径级的树木中,虽然株数很小,如最高碳贮量XIX级的树木株数仅为6株,是径级Ⅱ级 (7 cm≤DBH<9 cm)的1/5,但却占林分碳贮量的近1/3,这些大树木有很长的寿命,因此可以把它们视为长期的碳库;而青年林中最高碳贮量集中在中等径级,随着树木的生长,会在固碳方面起更大的作用,可视为潜在碳库.
3.3 乔木层碳贮量的树种间分配
无需考虑研究面积的大小,以树种个数与全部树种总数的商表示树种对林分的贡献情况,即相对多度=(树种的个数/样地全部树种的总数)×100%.相对碳贮量=(树种的碳贮量/样地全部树种的总碳贮量)×100%,可表示出树种对整个林分碳贮量的贡献情况.按照相对碳贮量大小,对2个林分树种进行编号与排序.
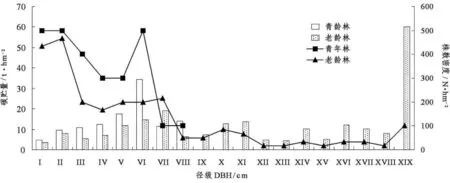
图2 不同林龄常绿阔叶林各径级碳贮量和株数Figure 2 DBH classes allocation of carbon storage and plant number in evergreen broad-leaved forests at different age
3.3.1 青年林碳贮量的树种间分配
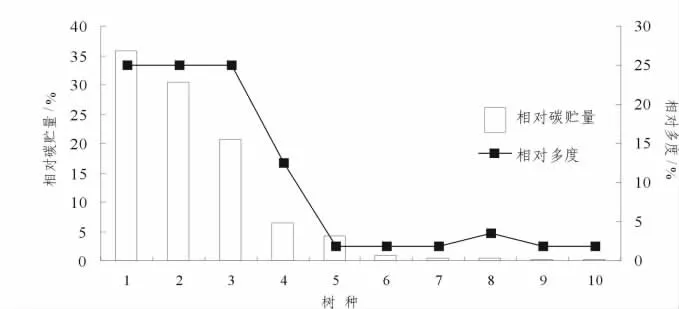
图3 青年林中不同树种的相对碳贮量和相对多度Figure 3 Relative carbon storage and relative abundance of different tree species in young forest
青年林中有10个树种 (图3),随着树种相对碳贮量的递减,相对多度呈现出不规则的变化趋势.在树种7(八角枫)、8(四照花)上出现不同,在相对碳贮量上八角枫为0.52%,四照花为0.50%,但两者的相对多度却是四照花>八角枫,从树种固碳能力上看:八角枫大于四照花.另外树种1(拟赤杨)、2(酸枣)、3(枫香)具有相同的相对多度,均为25%,但碳贮量却呈现出拟赤杨>酸枣>枫香,碳贮量分别为41.18 t·hm-2,34.99 t·hm-2,23.86 t·hm-2,对森林碳贮量的贡献出现差异,显示出三者固碳能力拟赤杨>酸枣>枫香.
3.3.2 老龄林碳贮量的树种间分配
老龄林中,同样按照碳贮量大小对树种进行编号与排序,即从1~33.
相对多度并没有随着相对碳贮量的递减而呈现递减的规律,而是呈现出波浪型的趋势 (图4).树种1(罗浮栲)相对丰度为13.38%,但具有最高的碳贮量,为82.71 t·hm-2,占总碳贮量的36.62%.而树种4(披针叶山矾)相对多度大于树种1,为15.27%,但碳贮量仅为15.5 t·hm-2,远远小于罗浮栲.这在一定程度上说明了优势树种不一定有高的碳贮量,对林分碳贮量的贡献也不一定是最高的.另外对各树种相对多度和相对碳贮量分析,可以看出各树种对林分碳贮量的贡献程度,对功能型结构最合适的描述则是相对碳贮量和相对多度[7].根据相对多度和相对碳贮量的大小将物种分为以下4类 (图5):少数/不重要 (0.1<相对多度≤1,0.1<相对碳贮量≤1);少数/重要 (0.1<相对多度≤1,10≤相对碳贮量≤100);丰富/不重要 (10≤相对多度≤100,0.1<相对碳贮量≤1);丰富/重要 (10≤相对多度≤100,10≤相对碳贮量≤100).
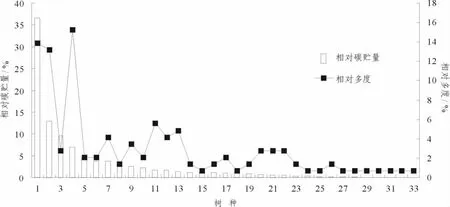
图4 老龄林中不同树种的相对碳贮量和相对多度Figure 4 Relative carbon storage and relative abundance of different tree species in old-growth forest
从图5可以看出,2个林分的大多树种处于中间状态,青年林没有少数/不重要的树种,却存在相对多度和相对碳贮量均高的树种;而老龄林存在少数/不重要、丰富/重要的树种.从水平方向上看,2个林分均为随着相对多度增加而呈相对碳贮量增加的趋势;垂直方向上看,相对多度一定,相对碳贮量也存在增加的趋势,这说明除了树种数量对碳贮量产生影响之外,树种的碳吸存能力 (表现为个体大小)对碳贮量也有影响.从树种整体水平上看,老龄林较青年林丰富.
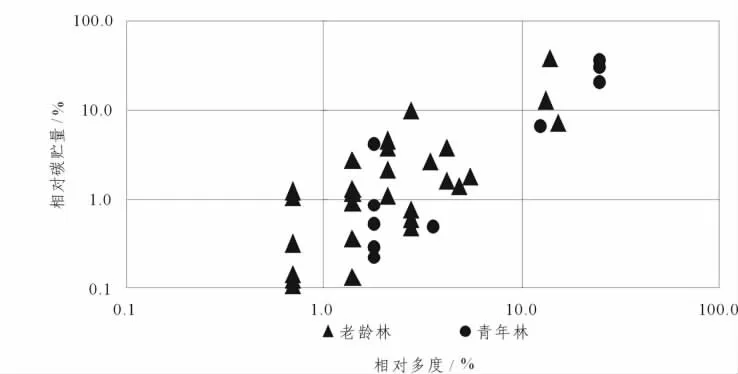
图5 不同林龄常绿阔叶林乔木层相对多度与相对碳贮量Figure 5 Relative abundance and relative carbon storage of arbor layer in evergreen broad-leaved forests at different age
4 讨论
根据估算公式得到青年林碳贮量为115.03 t·hm-2,老龄林碳贮量为224.43 t·hm-2,是青年林的2倍之多,这显示出碳贮量与林龄成正相关关系,与福建南平87年生杉木人工林乔木层碳库高于40年生杉木人工林这一研究相似[8],但后者差值不大,这可能与人为干扰等因素有关.且2个林分乔木层碳贮量大于福建省平均树干碳贮量 (73.43 t·hm-2),也高于福建南平楠木林乔木层碳贮量 (69.09 t·hm-2)[9].
本研究从每立木树高和胸径角度研究与碳贮量的关系.大多数研究都在于胸径径级碳贮量分配上,对于树高径级与碳贮量的研究甚少.在本研究中得出树高与碳贮量的偏离特征,即树高最高但碳贮量并不是最大.青年林树高径级Ⅶ级 (15 m≤H<17 m)有最高的碳贮量,而老龄林最高碳贮量则出现在Ⅷ级 (17 m≤H<19 m);而碳贮量的高度级分配与个体数的分布趋势不完全一致 (图1),这说明了碳贮量受胸径和树高的影响大于植株数量,唐旭利等在对鼎湖山季风常绿阔叶林碳贮量分布中发现类似的规律[10].青年林胸径径级Ⅵ (15 cm≤DBH<17 cm)出现最大碳贮量,而老龄林最大碳贮量出现在XIX级 (DBH≥41 cm),与鼎湖山山地常绿阔叶林、南亚热带常绿阔叶林对比[11],青年林与山地常绿阔叶林 (最大生物量出现在15~20 cm径级)相似,最大碳贮量均在<20 cm的小径级;而老龄林则与南亚热带常绿阔叶林 (最大生物量出现在35~50 cm径级)相似,大径级有最大碳贮量.从径级分布形态来看,2种林分碳贮量大体上符合递增的分布特征,与西双版纳原始热带季节雨林分布趋势较一致[12],但与四川云杉人工林符合倒钟型的生物量分布特征[13]、海南黎母山热带雨林的生物量径级分布呈递增分布[14]有较大差异.这说明碳贮量与林分每立木胸径有一定的关系,大树对森林碳贮量的贡献也是巨大的,Clark等在对哥斯达黎加的热带雨林生物量的研究中发现,大树对乔木层碳贮量的贡献为14%~30%[15];戚剑飞等对西双版纳石灰山季雨林的生物量及其分配规律研究中发现D为124 cm的大树其碳贮量占乔木层碳贮量的18.89%[16];而在本研究中老龄林DBH≥41 cm的树木仅有6株,仅为总株数的4.2%,但碳贮量却占总碳贮量的26.78%.
尽管树种对森林碳贮量的平均贡献基本上是与其相对多度一致,但是在对农林生态系统的研究中发现两者发生偏移[10].在本研究中也发现此情况,青年林中,拟赤杨、酸枣、枫香具有相同的相对多度,均为25%,但碳贮量却呈现出拟赤杨>酸枣>枫香;老龄林罗浮栲相对多度小于披叶针山矾,但对整个林分的碳贮量贡献上是罗浮栲远大于披叶针山矾.图5表明青年林和老龄林大多数树种对森林碳贮量贡献处于中间状态,青年林处于丰富/重要阶段的树种占总株数的25%,但对总碳贮量的贡献为30.5%;而老龄林处于丰富/重要阶段的树种占总株数的13.9%,但对总碳贮量的贡献为36.7%.Patricia等人对169种物种的地上碳贮量进行研究,仅仅13%的物种就对总碳贮量贡献90%,大多数物种对碳贮量的贡献很微小[7].在本研究中也发现同样的规律.
5 结论
青年和老龄常绿阔叶林乔木层碳贮量及株数的分配 (高度、径级、树种间)均呈现不一致的特征.株数集中在中低高度级、小径级,而碳贮量集中在大高度级、大径级;一般某一树种的多度最大,其碳贮量也最大,但不同树种对碳贮量的贡献仍受该树种个体碳吸存量的影响.林龄的不同导致青年和老龄常绿阔叶林碳贮量和株数分配 (高度、径级、树种间)不同.2个林分最大株数出现在Ⅲ高度级 (7 m≤H<9 m),但径级的分布则不一致;老龄林最大碳贮量所在的高度级和径级均大于青年林;老龄林树种碳贮量分配较青年林复杂,但2个林分碳贮量主要集中在少数树种上.
[1] Sabine C L,Heimann M,Artaxo P,et al.Current status and past trends of the global carbon cycle [C].In:Field C,Raupach M eds.The Global Carbon Cycle:Integrating Humans,Climate and the Natural World.Island Press,Washington,2004,17-44.
[2]IPCC(Intergovernmental Panel on Climate Change).Good Practice Guidance for Land Use,Land-Use Change and Forestry[R].IPCC/IGES,ISBN,Hayama,Japan,4288788200320,2003.
[3]Fornara D A,Tilman D.Plant functional composition influences rates of soil carbon and nitrogen accumulation[J].Journal of Ecology,2008,96:314-322.
[4]Henrya M,Tittonellc P,Manlaya R J,et al.Biodiversity,carbon stocks and sequestration potential in aboveground biomass in smallholder farming systems of western Kenya[J].Agriculture,Ecosystems& Environment,2009,129:238-252.
[5]Pan Hui,Huang Shi-de,Hong Wei,et al.The carbon storage and distribution in three Acacia plantations [J].Journal of Fujian College of Forestry,2009,29(1):28-32.[潘辉,黄石德,洪伟,等.3种相思人工林生态系统碳贮量及分配 [J].福建林学院学报,2009,29(1):28-32.]
[6]Wang Yi-xiang.Carbon stock of main forest types in Fujian province and carbon sequestration of Cunninghamis lanceolata plantation[D].Fujian Agriculture and Forestry University,2004.[王义祥.福建省主要森林类型碳库与杉木林碳吸存[D].福建农林大学硕士学位论文,2004.]
[7]Patricia B,Claire K,Miguel M R.Applying community structure analysis to ecosystem function:Examples from pollination and carbon storage[J].Ecological Applications,2005,15(1):360-375.
[8]Zhong Xian-fang,Yang Yu-sheng,Gao Ren,et al.Carbon storage and allocation in old-growth Cunninghamia lanceolata plantation in subtropical China[J].Journal of Subtropical Resources and Environment,2008,2(3):11-18.[钟羡芳,杨玉盛,高人,等.老龄杉木人工林生态系统碳库及分配 [J].亚热带资源与环境学报,2008,2(3):11-18.]
[9]Zheng Jin-xing,Liu Xiao-fei,Gao Ren,et al.Carbon storage and allocation in the Phoebe bournei plantations in Nanping,Fujian province[J].Journal of Subtropical Resources and Environment,2009,4(4):59-65.[郑金兴,刘小飞,高人,等.福建南平35 a生楠木林生态系统碳库及分配 [J].亚热带资源与环境学报,2009,4(4):59-65.]
[10]Tang Xu-li,Zhou Guo-yi,Wen Da-zhi,et al.Distribution of carbon storage in a lower subtropical monsoon evergreen broad:Leaved forest in Dinghushan Nature Reserve[J].Acta Ecologica Sinica,2003,23(1):90-97.[唐旭利,周国逸,温达志,等.鼎湖山南亚热带季风常绿阔叶林C贮量分布[J].生态学报,2003,23(1):90-97.]
[11]Liu Shen,Luo Yan,Huang Yu-hui,et al.Studies on the community biomass and its allocations of five forest types in Dinghushan Nature Reserve[J].Ecological Science,2007,26(5):387-393.[刘申,罗艳,黄钰辉,等.鼎湖山五种植被类型群落生物量及其径级分配特征 [J].生态科学,2007,26(5):387-393.]
[12] Zheng Zheng,Liu Hong-mao,Liu Lun-hu,et al.A study on biomass of the primary tropical seasonal rain forest in Xishuangbanna[J].Guihaia,1999,(19):309-314.[郑征,刘宏茂,刘伦辉,等.西双版纳原始热带季节雨林生物量研究 [J].广西植物,1999,(19):309-314.]
[13]Yan Wu-xian,Su Yi-ming,Liu Xing-liang,et al.Research on biomass and productivity of an artificial forest[J].Sichuan Research Institute of Forestry,1991,(12):17-22.[鄢武先,宿以明,刘兴良,等.云杉人工林生物量和生产力的研究 [J].四川林业科技,1991,(12):17-22.]
[14]Huang Quan,Li Yi-de,Lai Ju-zhang,et al.Study on biomass of tropical mountain rain forest in Limushan,Hainan Island[J].Acta Phytoecologica et Geobotanica Sinica,1991,(3):197-206.[黄全,李意德,赖巨章,等.黎母山热带山地雨林生物量研究[J].植物生态学与地植物学学报,1991,(3):197-206.]
[15]Clark D B,Clark D A.Landscape-scale variation in forest structure and biomass in a tropical rain forest[J].Forest Ecology and Management,2000,137:185-198.
[16]Qi Jian-fei,Tang Jian-wei.Biomass and its allocation pattern of monsoon rain forest over limestone in Xishuangbanna of Southwest China[J].Chinese Journal of Ecology,2008,27(2):167-177.[戚剑飞,唐建维.西双版纳石灰山季雨林的生物量及其分配规律 [J].生态学杂志,2008,27(2):167-177.]
Allocation of Carbon Storage in the Arbor Layer of Young and Old-growth Evergreen Broad-leaved Forests in Wanmulin
HUANG Rong,WANG Chao,YANG Zhi-jie,CHEN Guang-shui
(1.Institute of Geography,Fujian Normal University,350007,Fuzhou,China;2.State Key Laboratory of Mountain Ecology in Humid Subtropics,350007,Fuzhou,China)
In order to investigate the influence of forest biomass carbon on forest carbon storage,carbon sink measurement and management,as well as biodiversity conservation,this paper analyses the evergreen broad-leaved forests,including the young forest(about 47 a)and old-growth forest(about 120 a)in the Wanmulin Nature Reserve in Fujian.According to the results of biodiversity investigation and Biomass Different Speed Equation,we figure out the carbon storage in tree layer and its distribution features in different heights,diameter classes and tree species.The results are shown as follows:1)The carbon storage of young forest is 115.03 t·hm-2,while that of the old-growth forest is 224.43 t·hm-2.2)In young forest,the maximum carbon storage is 23.17 t·hm-2in height classⅦ (15~17 m),in old-growth forest,it(62.96 t·hm-2)happens in height classⅧ (17 ~19 m).3)In young forest,the maximum carbon storage which amounts to 34.25 t·hm-2,lies in DBH classⅥ (15 cm≤DBH<17 cm);in old-growth forest,the maximum carbon storage lies in class XIX(DBH≥41 cm)which amounts to 60.03 t·hm-2,taking up 26.78%of total carbon storage.4)Most carbon is stored in a small minority of trees,in other words,most tree species con-tribute only a tiny part to the total carbon storage.
arbor carbon storage;carbon allocation;broad-leaved evergreen forests;abundance;richness
S718
A
1673-7105(2011)02-0029-07
2011-03-15
973计划前期研究专项课题 (2010CB434804)、福建省海西杰出青年人才基金项目 (2010J06009)
黄蓉 (1987—),女,江西赣州人,硕士研究生,从事森林碳循环研究,(E-mail)hr_009@163.com.
陈光水 (1976—),男,福建仙游人,副教授,博士,硕士研究生导师,从事根系生态学和森林碳循环研究,(E-mail)gshuichen@163.com.
猜你喜欢
农业科技与信息(2022年9期)2022-06-18
现代农业科技(2020年11期)2020-06-21
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
农家科技下旬刊(2017年8期)2017-11-13
江苏农业科学(2017年6期)2017-05-11
绿色科技(2017年3期)2017-03-14
科技创新导报(2016年21期)2016-12-17
学苑创造·B版(2015年12期)2016-06-23
江苏农业科学(2014年11期)2015-01-15
