
广东省西江与东江草鱼群体遗传多样性的RAPD分析
2011-08-30 06:11黎卓键
科技传播 2011年17期
黎卓键,刘 丽
华南农业大学动物科学学院,广东广州 510642
草鱼(Ctenopharyngodon idellus)俗称草鲩,英文名为Grass carp,系属脊椎动物门(Vertebrata)、硬骨鱼纲(Osteichthys)、鲤 形 目(Cypriniformes)、 鲤 科(Cyprinidae)、 雅 罗 鱼 亚 科(Leuciscinae)、草鱼属(Ctenopharyngodon Steindachner,1866)[1]。草鱼作为四大家鱼之首,其所形成的产业对广东省淡水渔业乃至全国的淡水渔业的发展都起到了极其重要作用。
随 机 扩 增 多 态 DNA(random amplified polymorphic DNA,RAPD)是建立在PCR技术基础上检测DNA序列多态性和建立分子遗传标记的技术,Williams等[2]和Welsh等[3]运用随机引物扩增寻找多态DNA片段作为分子标记。如今RAPD技术在生物的遗传多样性、群体遗传学、分类学及农牧业的遗传育种等研究中得到广泛应用[4]。本研究旨在用RAPD技术对广东草鱼进行群体的亲缘关系探讨,以期为了解广东省草鱼资源现状提供相关的遗传背景资料和理论依据。
1 材料和方法
1.1 材料
2008年5 月~2008年7月于西江肇庆江段、东江新丰江水库和佛山龙江镇锦记鱼苗场共取90尾草鱼个体的背部肌肉放在冻存管置于液氮罐,运回实验室液氮保存。采集点分布见图1,采集信息见表1。

图1 样品采集地点
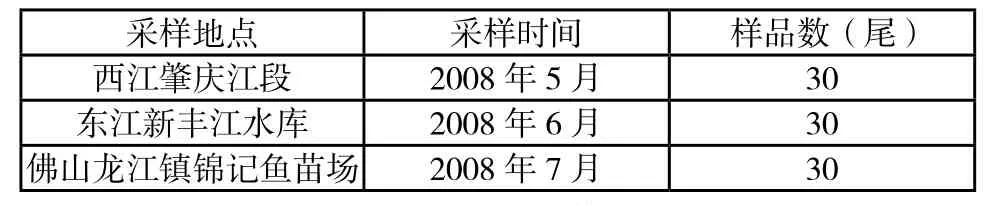
表1 草鱼采样信息
1.2 DNA提取
取液氮保存肌肉组织大约10mg,与200μl细胞裂解液混匀,加入1.5μl蛋白酶K[5],置于55℃水浴锅中消化过夜。从水浴锅中取出,加入300μl三氯甲烷混匀,上下摇动10min~15 min。10 000×g离心2min,取约200μl上清液,置于1.5 ml心管中。在离心管中加入250μl沉淀液,混匀,室温放置2 min,10 000×g离心2min。倒掉上清液,立即加入50μl 2M NaCl,轻轻振荡直至DNA样品完全溶解,加入1.5μl RNase A,然后置于37℃水浴锅中水浴5min~10min。加入150μl冷乙醇,-20℃放置10min,10 000×g离心4 min,倒掉乙醇,加入300μl 75%乙醇清洗,10 000×g离心2 min,倒掉75%乙醇,自然风干至无酒精味,加入50μl TE,37℃水浴锅中水浴溶解,4℃保存备用。
1.3 随机引物及筛选
从已研究草鱼的RAPD文献中选取60条引物,以高纯度的草鱼基因组DNA为模板,对60个引物进行筛选,从中选取扩增条带多且清晰的7个随机引物进行PCR扩增。
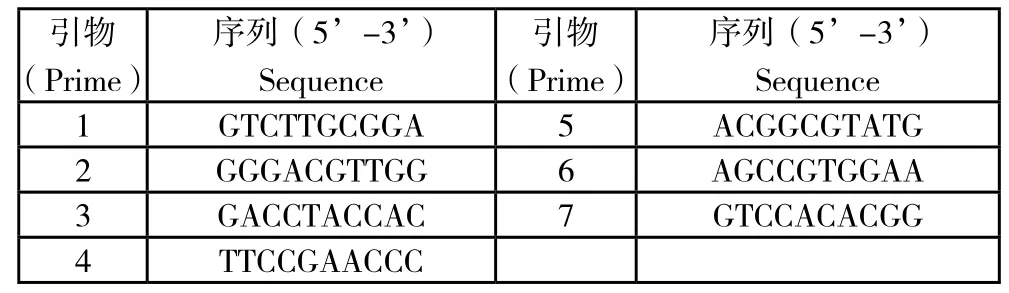
表2 7个引物的名称和序列
1.4 RAPD-PCR“标准”反应体系
扩增条件为: 94℃预变性5min,94℃变性1min,36℃退火1min,72℃延伸2min,40个循环,72℃延伸10min,4℃保存。产物在1.5%琼脂糖凝胶上,用恒压80V电泳1小时,溴化乙锭染色,紫外透射仪下观察并照相。根据 50bp DNA ladder(takara)对扩增的片段进行确认。
1.5 数据分析
1.5.1 RAPD片段的观察与统计
根据PCR产物的电泳带型,进行群体内和群体间的比较。统计时仅记录清晰、稳定的电泳带。在同一个引物相同的迁移率上,不同个体间出现的带记为1,未出现的带记为0,在excel表中按格式输入每一个引物的0、1距阵。
1.5.2 多态位点比例
利用扩增片段原始数据的0-1矩阵,使用RAPD data analyzer 1.3v数据分析软件,计算群体内个体间的多态位点数和总位点数,然后根据如下公式计算多态位点比例:多态位点比例(P)=多态位点数/总位点数×100%
1.5.3 遗传相似度和遗传距离
根据统计的原0-1矩阵,进行个体间与群体间的比较分析。按公式Sxy=2nxy/(nx+ny)计算出个体间遗传相似性指数(nxy是个体x和y的共有带数,nx和ny分别是个体x和y扩增的扩增带数)。
群体间的相似性指数(Sij)为种群i中的个体和种j中的个体随机组合所得的相似指数和平均值[6-7]。用公式D=1-S计算个体间及种间的遗传距离。最后用MEGA2. 1软件的类平均聚类法(unweighed pair-groupmethod of analysis, UPGMA) 和 邻 接 法(neighbor joining, NJ)对0-1矩阵值进行聚类分析。
1.5.4 Shannon遗传多样性指数
种群内的遗传分化可用Shannon遗传多样性指数进行估算,指数越大遗传多样性越大,种群分化的程度越高。每个RAPD多态位点上,可扩增出的带出现频率是p(出现次数/个体总数),不能扩增出的带频率是q,其中p+q=1。根据RAPD各多态位点的基因型频率,应用下列公式计算Shannon遗传多样性指数指数 H,从而估算种群内的遗传距离。用公式:H =-∑ pi Ln pi进行计算,即每个RAPD多态位点上带出现的频率p,取Ln为自然对数。∑是每个多态位点的值求和[8]。
2 结果
2.1 模板DNA的检测

图2 草鱼群体基因组DNA琼脂糖凝胶(0.7%)电泳图
本实验提取得到的DNA,经0.7%的琼脂糖凝胶电泳显示DNA带型致密,分子量较大,约在20kb左右,符合RAPD对模板DNA质量的要求。图2显示的是草鱼群体基因组DNA琼脂糖凝胶(0.7%)电泳图
2.2 RAPD扩增结果
在选用的60个随机引物中,有53个引物无扩增产物或因结果不稳定无法做进一步分析。其余7个随机引物在3个群体的90个个体中扩增结果稳定、重复性佳,共在西江草鱼群体中检测到62个位点、在东江草鱼群体中检测到59个位点、在良种场草鱼群体中检测到56个位点,检测到的平均位点数分别为西江8.86,东江8.43,良种场8个,得出3个草鱼群体的多态位点比例分别为21.00%,18.64%,33.96%。其多态位点比例的大小顺序为:良种场草鱼〉西江草鱼〉东江草鱼。
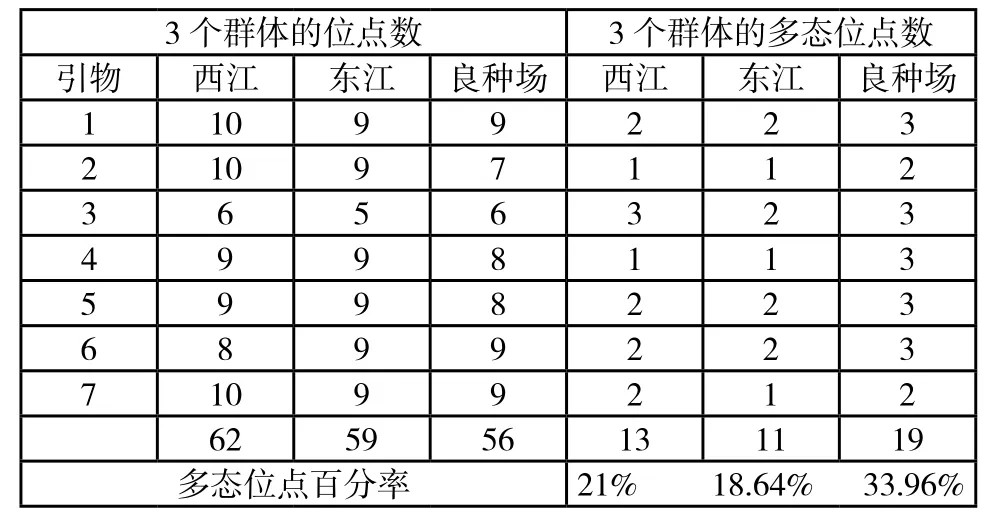
表3 RAPD扩增的位点数、多态位点数以及多态位点比例

引物5在西江草鱼群体的RAPD扩增物琼脂糖凝胶(1.5%)电泳图
注:1.50bp分子量标准;2~16.引物5在西江草鱼群体的扩增条带箭头所示西江和东江草鱼群体中检测到而良种场草鱼群体中没有检测到的特异谱带(约200bp处)

引物5在东江草鱼群体的RAPD扩增物琼脂糖凝胶(1.5%)电泳图
注:1.50bp分子量标准;2~15.引物5在东江草鱼群体的扩增条带箭头所示西江和东江草鱼群体中检测到而良种场草鱼群体中没有检测到的特异谱带(约200bp处)
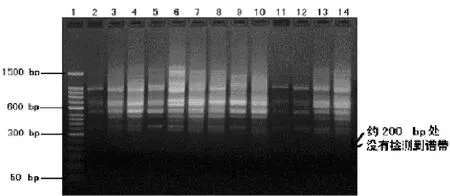
引物5在良种场草鱼群体的RAPD扩增物琼脂糖凝胶(1.5%)电泳图
注:1. 50bp分子量标准;2~14. 引物5在良种场草鱼群体的扩增条带良种场草鱼群体中在约200bp处没有检测到相应的特异谱带
图3 引物5在3种鱼群体基因组的RAPD-PCR扩增图谱
2.3 3个草鱼群体的群体内遗传相似度和遗传距离
运用NTSYS-pc 2.2软件计算3个草鱼群体内的遗传相似度和遗传距离,结果见表4。由表4中可以看出遗传相似系数在0.8863~0.9811,平均值为0.9435。这3个草鱼群体内的遗传距离大小顺序恰恰相反,其大小顺序为良种场草鱼群体〉西江草鱼群体〉东江草鱼群体

表4 三个草鱼群体内的遗传相似度和遗传距离
2.4 Shannon遗传多样性指数
用Shannon遗传多样性指数计算了3个草鱼群体的群体内遗传多样性,其结果见表5。由于不同的随机引物检测到的位点数及多态性位点数均有所不同,按各个引物计算的遗传多样性指数值就有一定的差异,因此计算各个引物的Shannon遗传多样性指数后求出其平均值。

表5 三个草鱼群体的群体内的Shannon遗传多样性指数
Shannon遗传多样性指数越大表明遗传多样性越大,种群分化的程度越高。数据结果表明:良种场的草鱼群体遗传多样性〉西江草鱼群体遗传多样性〉东江草鱼群体遗传多样性。其结果和表4计算所得3个草鱼群体内的遗传距离的结果是相吻合的。
2.5 3个草鱼群体间的遗传相似度和遗传距离
通过对RAPD谱带的比较和分析,分别计算出草鱼3个群体间的遗传相似性指数和遗传距离,结果见表6。

表6 草鱼三个群体间的遗传相似性指数和遗传距离
如表6所示,群体间遗传相似性指数西江和东江之间最大,为0.9454;东江和良种场次之,为0.8821;西江和良种场之间最低,为0.8532。群体间的遗传距离良种场和西江之间最大,为0.1468;良种场和东江之间次之,为0.1179;西江和东江之间最小,为0.0546。草鱼群体间的平均遗传距为0.1064,并根据表的遗传距离,构建聚类图(图4)。西江、东江草鱼群体先聚为一类,然后再与良种场草鱼群体相聚。

图4 3个草鱼群体遗传距离聚类图
3 讨论
聚类分析可以对物种的亲缘疏远关系进行比较,量化种群间的差异程度。形态距离的聚类分析结果表明,西江和东江之间的距离最近,形态也很相近,良种场群体和西江、东江这两个群体之间在形态上有一定差异。鱼类形态特征很大程度上取决于遗传特性。而鱼类形态学性状也与鱼类生活成长环境条件相紧密联系的。形态性状的稳定与演化,都是鱼类对特定栖息环境条件的长期适应,在生活习性、形态结构和生理机能等方面发生相适应的变化。形态性状是与生理机能、生活习性的变化相一致的。综合分析结果表明:在形态方面,3个草鱼群体内差异不大,而东江和西江草鱼群体间差异不大,这两个地方群体与良种场群体间有一定差异,这同它们来源差异是相符合的,也可能与遗传背景有关。要从形态上对不同地方草鱼进行区分不能要求只用少数指标就能鉴别,而要用多项参数综合判别。本实验对3个草鱼群体在分子水平进行种质鉴定研究是为了尽最大可能研究不同地方草鱼遗传变异的水平;研究草鱼物种的进化潜力;生物资源的永续利用,为草鱼资源的生产利用提供依据。
由于新丰江水库是90世纪60年代苏联援建,因此建库已超过50年。结果导致新丰江水库的野生草鱼属于一个比较密闭的群体,它和西江野生的草鱼群体在形态分析上的相似性比较大,而这两个地方的草鱼与良种场的草鱼在形态上有一定的差异。这与所采良种场的草鱼是来自长江水系的是相吻合的。因为在珠江水系基本上没有草鱼产卵场,草鱼的产卵场基本在长江中游[9]。
鱼类物种是以种群的形式而存在的,不是个体的随意综合,而是分别结合成或大或小的种群,种群之间并不是以连续的方式存在,组成一个统一的繁殖群体。在逐渐形成一个稳定群体的过程中,鱼类种群由于突变后的自然选择而出现变异和进化。鱼类种群的遗传变异水平是衡量鱼类资源状况和遗传背景的一个重要标准。通过研究掌握现有鱼类资源的遗传多样性,对不同生物群体进行鉴别,从而可以清楚地掌握种群的分布;可以知道各个群体之间的亲缘关系。遗传多样性的降低将导致其适应能力降低,有害隐性基因增加及经济性状衰退,会产生严重的遗传瓶颈效应,最后导致物种退化。因此,检测种群的遗传变异和多样性水平是十分必要的。迄今为止,已有多种不同的检测手段检测遗传变异水平,RAPD分子标记技术由于其操作简便,产生的多态性丰富等优点,可以直接检测基因组中的变异情况,为种群和种群内个体之间的遗传多样性研究提供有力的手段
目前己有不少学者应用RAPD技术对草鱼进行了遗传多样性分析[10-11]。薛国雄等[10]对长江、珠江、黑龙江三江水系的草鱼种群进行RAPD分析结果表明:每一水系的草鱼种群均有其特征性基因图谱,可作为种群鉴定的依据。章怀云[12]等对草鱼和红鲤进行了RAPD分析,结果表明草鱼的遗传多样性明显低于红鲤。目前在草鱼的多样性研究中,不同方法得到的研究结果均显示草鱼的遗传多样性不丰富[11-13]。张四明[11]采用RAPD技术对长江中游的湖北嘉鱼与江西瑞昌两个地理群体和中游的汉江和湘江两大水系的鲢鱼和草鱼的群体进行了遗传学研究。发现草鱼遗传多样性从大到小的分布是瑞昌、汉江、湘江、嘉鱼,四个地理群体草鱼的遗传分化度较低,可能与地理位置较近和基因交流频繁有关;草鱼有可能在历史上经历了“遗传瓶颈”效应,推测草鱼很有可能是一个遗传多样性贫乏的常见物种[14]。
本实验中,利用7个引物对珠江水系西江和东江草鱼群体进行RAPD分析,并以良种场草鱼群体作为对照,得到的结果为:草鱼群体间的遗传距离西江和良种场之间最大,为0.1468;东江和良种场之间次之,为0.1179;西江和东江之间最小,为0.0546。3个草鱼群体内的遗传差异很小,草鱼的遗传多样性很低,和已有的研究结果相符。今后研究过程中,应采用不同方法(形态学、细胞遗传学、生物化学、分子生物学等技术),从不同层次上(群体、个体、细胞、分子水平)进行研究,并且进行综合分析,使人们对所研究的对象有一个更加全面准确的认识,以便更好的利用草鱼鱼类资源。
[1]郑慈英.珠江鱼类志[M].科学出版社,1989:77-78.
[2]WILLIAMS J G,KUBELIK A R,LIVAK K J.DNA Polymorphisms Amplified by Arbitrary Primers are Useful as Genetic Markers[J].NuclAcidsRes,1990,18:6531-6535.
[3]WELSH J,PETERSON C,MC CLELLAND M.Polymorphisms Generated by Arbitrarily Primed PCR in Mouse:Application to Strain Identification and Genetic Mapping[J].NuclAcidsRes,1991,20:303-306.
[4]HUANG JC,WANGW K,HONG KH,et al.Population Differentiation and Phylogeography of Hygrophila PogonocalyxBased on RAPDs Fingerprints[J].Aquat Bot,2001,70(4):269-380.
[5]萨姆布鲁克.J,弗里奇.EF,曼尼阿蒂斯.T.分子克隆实验指南[M].2版.北京:科学出版社,2003,1992,461-471.
[6]LYNCHM.The Similarity Index and DNA Fingerprinting[J].MolBiolEvo,l 1990,7:478-484.
[7]BARDAKAI F,SKIBINSKID O F.Application of the RAPD Technique in theTilapia Fish: Species and Subspecies Indent fication [J].Heredity,1994,73:117-123.
[8]Wachira F.N,Waugh R,Powell W,Hackett C.A.Detection on genetic diversity in tea (Camellia sinensis)using RAPD markers.Genome,1995,38:201-210.
[9]王尚玉,廖文根,陈大庆,段辛斌,彭期冬,王珂,李翀.长江中游四大家鱼产卵场的生态水文特性分析[J].长江流域资源与环境,2008,17(6):892-897.
[10]薛国雄,刘棘,刘洁.三江水系草鱼种群RAPD分析[J].中国水产科学,1998,5(1):1-5.
[11]张四明,汪登强,邓怀.长江水系鲢和草鱼遗传结构及变异性的RAPD研究[J].水生生物学报,2001,25(4):324-330.
[12]章怀云,刘荣宗,张学文.草鱼和鲤群体遗传变异的RAPD指纹分析[J].水生生物学报,1998,22(2):168-173.
[13]廖小林,俞小牧,谭德清,童金苟.长江水系草鱼遗传多样性的微卫星DNA分析[J].水生生物学报,2005,29(2):113-119.
[14]谭书贞.应用DNA和同工酶遗传标记检测长江3个草鱼群体的遗传多样性[D].天津师范大学,2007:37-39.
猜你喜欢
中国水产(2023年8期)2023-09-09
系统工程学报(2021年4期)2021-12-21
浙江柑橘(2020年3期)2020-10-12
新世纪智能(高一语文)(2020年12期)2020-06-01
岭南音乐(2016年4期)2016-11-21
浙江柑橘(2016年4期)2016-03-11
浙江柑橘(2016年3期)2016-03-11
浙江柑橘(2016年2期)2016-03-11
——东江鱼(实业)集团有限公司
湖南农业(2015年10期)2015-03-18
中国医药导报(2015年27期)2015-02-28
