
抗高效氟吡甲禾灵日本看麦娘生长速率与生长密度研究
2011-08-16 05:14:48董立尧
杂草学报 2011年2期
张 彬,李 俊,2,董立尧,2
(1.南京农业大学植物保护学院/农业部作物病虫害监测与防控重点开放实验室,江苏南京 210095; 2.南京农业大学理学院/江苏省农药学重点实验室,江苏南京 210095)
抗高效氟吡甲禾灵日本看麦娘生长速率与生长密度研究
张 彬1,李 俊1,2,董立尧1,2
(1.南京农业大学植物保护学院/农业部作物病虫害监测与防控重点开放实验室,江苏南京 210095; 2.南京农业大学理学院/江苏省农药学重点实验室,江苏南京 210095)
抗性日本看麦娘种群与敏感种群种子在低温(4 ℃)清水浸泡处理超过6 d后萌发率均能达90%以上,但抗性种群休眠解除的时间要长于敏感种群;敏感种群在播种后的前15 d内生长速率快于抗性种群,但此后抗性种群生长速率逐渐超过敏感种群;抗性种群在播种后20 d开始长出第2片叶,比敏感种群晚2 d,播种后25 d两种群出现第2片叶的比例均超过95%,进一步说明在生长后期抗性种群生长速率超过敏感种群;在相同的播种密度下,抗性种群长势优于敏感种群,150~200株/0.11m2的密度范围内有利于日本看麦娘的生长,抗性种群的最佳生长密度为208株/0.11m2,而敏感种群为186株/0.11m2。
日本看麦娘; 解除休眠; 生长速率; 生长密度
种子休眠是大多数杂草种子都具有的特性,是杂草在长期自然选择过程中形成的对不良环境的适应[1]。韩建国将种子休眠定义为“具有生活力而停留在不能发芽的状态”[2]。而国外学者则将种子休眠定义为“成熟而完好无损的种子在最适宜的条件下不能发芽或延迟发芽的现象”[3-4]。种子休眠对处于恶劣环境中的植物保持其自身繁衍发展具有重要的生态学意义[5]。大多杂草种子都具有休眠的特性,不同种类的植物及不同品种之间种子休眠时间长短差异很大。杨彩宏等分别采用地下沉积和低温清水浸泡两种休眠解除方法对日本看麦娘种子休眠解除进行了相关的研究,结果表明低温清水浸泡法能有效的解除所有日本看麦娘种群种子的休眠[6]。适合度(fitness)简单的说就是植物的存活及繁殖成效,即在某种环境条件下,某已知基因型的个体将其基因传递到后代基因库中的相对能力,是衡量个体存活和繁殖机会的尺度[7-8]。适合度越大,存活和繁殖机会越高。国内外对抗性种群适合度的研究已经有很多报道,大多学者已经证明,新的抗性种群的产生会伴随着适合度的降低[9-11],但也有研究表明抗药性能导致其适合度升高[12-13],因此对两种杂草种群之间相对适合度的比较研究就显得非常重要。
1 材料与方法
1.1 供试材料
日本看麦娘(AlopecurusjaponicusSteud.)种子采集于2009年6月份。通过种子生物测定法检测不同地点日本看麦娘种群的抗性水平。经测定,将采集于江苏省句容市油菜田的日本看麦娘作为抗性种群,将江苏省南京市紫金山草坪地中的日本看麦娘作为敏感种群(种子萌发率均为90%以上)。
供试除草剂为10.8%高效氟吡甲禾灵EC,美国陶氏益农公司生产。
1.2 试验方法
1.2.1 低温清水浸泡时间对解除抗高效氟吡甲禾灵日本看麦娘种子休眠的方法研究 取足量大小均匀、籽粒饱满的成熟日本看麦娘种子置于洁净的培养皿中,加适量的蒸馏水(以浸泡种子为准),置于4 ℃冰箱中。分别在第0、1、2、3、4、5、6、7、8、9天取出40粒放于直径为9 cm铺有1张滤纸的培养皿中,加5 mL超纯水,加盖以保持水分,置于光照培养箱中培养观察(温度:白天18 ℃,夜间13 ℃;光周期:12 D/12 L),4次重复。每天统计种子的萌发数并计算萌发率(胚芽长至2 mm时即为萌发),并比较两个种群种子萌发率间的差异。
1.2.2 抗高效氟吡甲禾灵日本看麦娘幼苗生长速率的研究 试验采用盆钵法。将细土均匀的装入直径为9 cm的塑料盆钵内,每盆播30粒已解除休眠的日本看麦娘种子,放于室内培养箱中培养,培养条件如1.2.1。各盆钵土壤始终保持湿润,4次重复。出苗后每盆定苗20株,每隔2 d记录日本看麦娘的株高,观察记录杂草长第2片叶的时间,并比较两个种群幼苗的生长速率。试验期间人工拔除其他杂草。
1.2.3 抗高效氟吡甲禾灵日本看麦娘最佳种群生长密度的研究 试验采用盆钵法。将均匀的细土装入直径为11.5 cm的塑料盆钵内,每盆分别播50、40、30、20、10粒已解除休眠的日本看麦娘种子,放于室内培养箱中培养,培养条件如1.2.1。各盆钵土壤始终保持湿润,4次重复。出苗后分别每盆定苗40、35、30、25、20、15、10、5株,出苗后第35天测量日本看麦娘的株高,并比较两个种群间幼苗株高的差异。试验期间人工拔除其他杂草。

其中,x为每盆的杂草株数,R为盆钵的半径。
1.3 统计分析
试验采用Excel 2003计算平均值和标准差并作图,用DPS v3.01统计软件进行统计分析。
杂草随时间变化而变化规律应用一元回归模型进行研究,即以杂草生长时间为自变量(x),以杂草的株高为因变量(y),公式为y=f(x)。以相关系数最大、离回归方差最小为原则进行优选[14-15]。
不同种植密度下日本看麦娘的长势应用一元二次曲线方程模型进行研究,即以杂草的种植密度为自变量(x),以杂草的株高为因变量(y),公式为y=ax2+bx+c。以相关系数最大、离回归方差最小为原则进行优选[14-15]。
2 结果与分析
2.1 低温清水浸泡时间对解除抗高效氟吡甲禾灵日本看麦娘种子休眠的影响
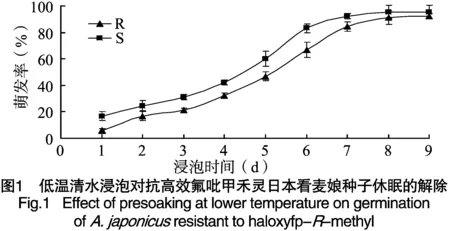
抗高效氟吡甲禾灵日本看麦娘种子休眠解除时间长于敏感种群(图1)。随着低温清水浸泡时间的延长,2个日本看麦娘种群种子萌发率均逐渐增加。未经低温清水浸泡处理的两个日本看麦娘种群种子萌发率很低,分别只有5.56%、16.67%。在第6天时2个日本看麦娘种群种子萌发率分别达到84.44%、92.22%,但抗性种群萌发率仍低于敏感种群。因此,抗性日本看麦娘种群种子比敏感种群难于解除休眠。
2.2 抗高效氟吡甲禾灵日本看麦娘生长速率
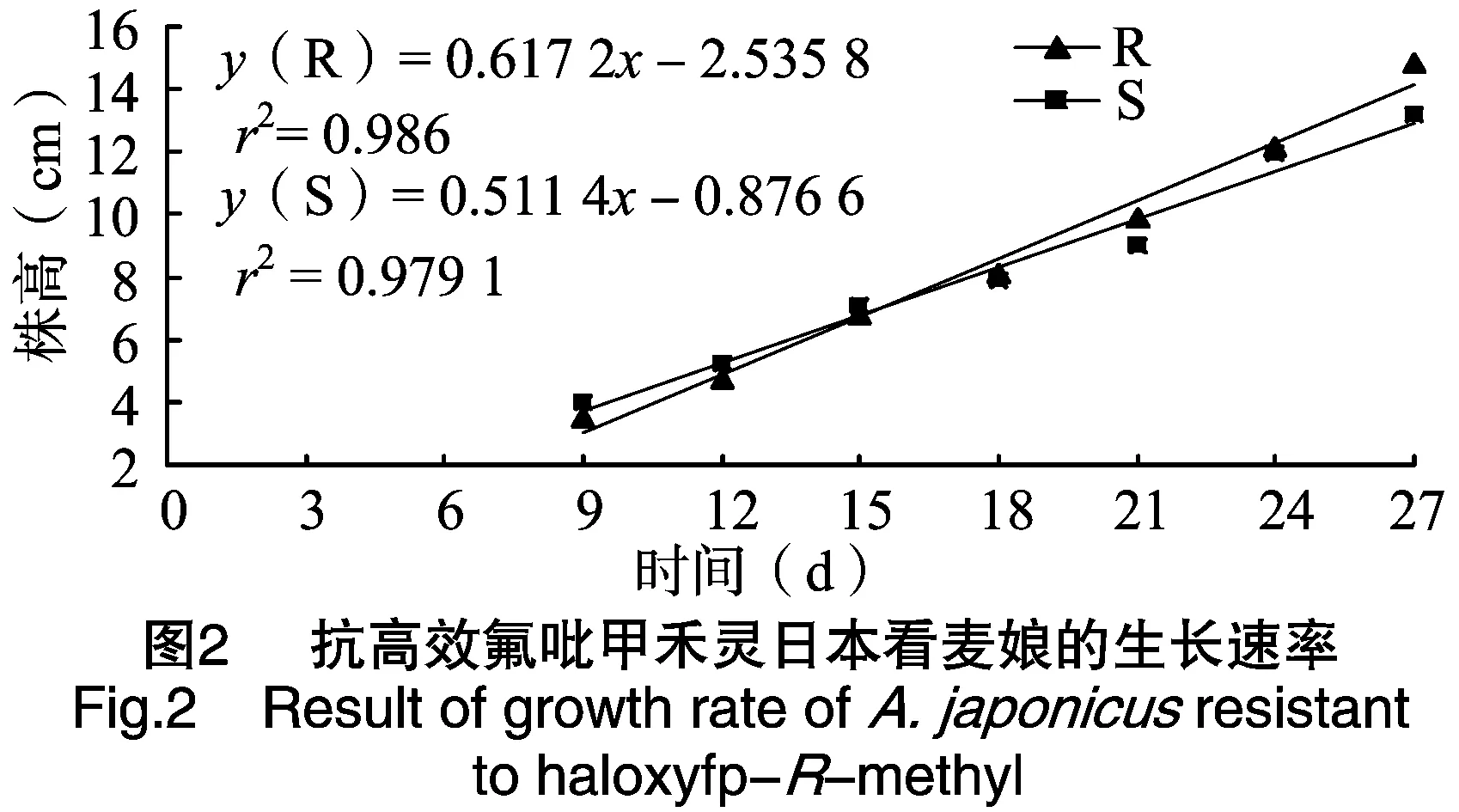
抗高效氟吡甲禾灵日本看麦娘生长速率不同于敏感种群(图2)。在生长早期,抗性种群生长速度稍微慢于敏感种群,但生长18 d以后,抗性种群生长速率逐步加快,并超过敏感种群。通过两个日本看麦娘种群生长速率的线性比较也验证了这点(R为0.617 2;S为0.511 4)。
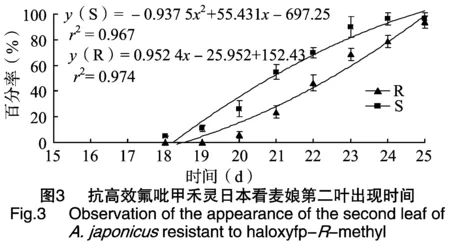
抗高效氟吡甲禾灵日本看麦娘第二片叶片的出现时间晚于敏感种群(图3),在种植第18天时,敏感种群生出第二片叶,而第19天时抗性种群开始生第二片叶,第25天时,两种日本看麦娘种群几乎都长出第二片叶,出现比率达到95%以上。通过曲线回归方程也可以看出抗性种群的二次项系数为0.952 4,敏感种群的二次项系数为-0.937 5,说明抗性日本看麦娘种群在生长后期快于敏感种群。
2.3 抗高效氟吡甲禾灵日本看麦娘最佳种群生长密度
在同等密度下抗性日本看麦娘种群长势优于敏感种群(图4)。150 ~200株/0.11m2种植密度有利于日本看麦娘的生长,低密度和高密度都不利于日本看麦娘的生长。初步建立了日本看麦娘种群密度与株高的二次曲线方程,通过曲线方程可以推算出抗性日本看麦娘种群的最佳生长密度为208株/0.11m2,大于敏感种群的186株/0.11m2。

3 讨论
种子休眠是植物在长期系统发育过程中形成的抵抗不良环境条件的适应性,是调节种子萌发的最佳时间和空间分布的有效方法,具有普遍的生态意义。种子休眠时间的长短因不同种类和不同品种间差异很大,而且不同种子破除休眠的方法不尽相同。对于室内杂草的研究,低温清水浸泡破除日本看麦娘种子休眠是最有效、最快捷的方法[6]。试验中通过对抗高效氟吡甲禾灵日本看麦娘种群种子低温清水浸泡解除休眠研究发现,与敏感种群相比,抗性种群的休眠解除明显困难。但到目前为止,出现这种结果的原因尚未有相关的报道和说明,因此需要进一步研究。
大多抗性种群的产生都会伴随其适合度的降低,但本试验研究发现抗性日本看麦娘种群在某些特性上优于敏感种群。在生长早期,抗高效氟吡甲禾灵日本看麦娘种群生长速率慢于敏感种群,但生长后期抗性种群生长速率及长势要优于敏感种群,与Sibony and Rubin(2002)和Holt and Thill(1994)报道的致使产生抗性的变异并没有降低适合度相吻合。Shrestha等在2010报道了对草甘膦产生抗性的加拿大乍蓬(Conyzacanadensis)在田间试验研究中表现出强于敏感种群的竞争优势[16],但对于抗性种群适合度增高的原因尚未见报道,尤其是本试验中发现的抗性日本看麦娘种群其表现出的适合度增高的原因,需要进一步的试验证明。
日本看麦娘属群集性生长的一种杂草,田间出苗率很高[17]。过高或过低的种群密度都不利于其正常的生长。试验证明,150~200株/0.11m2范围内有利于日本看麦娘的生长,抗性日本看麦娘种群的最佳生长密度208株/0.11m2,而敏感型为186株/0.11m2,这是否与抗性和敏感两种种群的生长环境有关,需要进一步的试验研究。但其最佳种群密度结果的发现,对以后田间日本看麦娘的控制和防除有一定的帮助和支持。
[1]苏少泉. 杂草科学[M]. 北京:农业出版社,1993:24-32.
[2 ]韩建国,李青丰,陈宝书,等. 牧草种子学[M]. 北京:中国农业大学出版社,2000:68-82.
[3]Prada D,Romagosa I,Ullrich S E,et al. A centromeric region on chromosome 6(6H) affects dormancy in an induced mutant in barley[J]. Journal of Experimental Botany,2005,56(409):47-54.
[4]Romagosa I,Prada D,Moralejo M A,et al. Dormancy,ABA content and sensitivity of a barley mutant to ABA application during seed development and after ripening[J]. Jourmal of Experimental Botany,2001,52(360):1499-1506.
[5]张 鹏,沈海龙. 白蜡树属树种种子休眠及其萌发的调控[J]. 植物生理学通讯,2006,42(2):354-358.
[6]杨彩宏,董立尧,李 俊,等. 油菜田日本看麦娘对高效氟吡甲禾灵抗药性的研究[J]. 中国农业科学,2007,40(12):2759-2765.
[7]刘凤沂,须志平,薄仙萍,等. 昆虫抗药性与适合度[J]. 昆虫知识,2008,45(3):374-378.
[8]孟香清,芮昌辉,赵建周,等. 抗三氟氯氰菊酯棉铃虫种群相对适合度研究[J]. 植物保护,1998,24(6):12-14.
[9]Park K W,Smith C A,Ball D A,et al. Ecological fitness of acetolactate synthaseinhibitor-resistant and -susceptible downy brome (Bromustectorum) biotypes[J]. Weed Science,2004,52:768-773.
[10]Hartzler R G,Battles B A. Reduced fitness of velvetleaf (Abutilontheophrasti) surviving glyphosate[J]. Weed Technology,2001,15:492-496.
[11]Massinga R A,Al-Khatib K,Amand P St,et al. Relative fitness of imazamox-resistant common sunflower and prairie sunflower[J]. Weed Science,2005,53:166-174.
[12]Steckel L E,Gwathmey C O. Glyphosate-resistant horseweed (Conyzacanadensis) growth,seed production,and interference in cotton[J]. Weed Science,2009,57:346-350.
[13]Poston D H,Wilson H P. Growth and development of imidazolinone-resistant and -susceptible smooth pigweed biotypes[J]. Weed Science,2002,50:485-493.
[14]王顺建,张 玉,株良备. 安徽淮北地区麦田蚤缀生物学特性及生态经济阈值研究[J]. 杂草科学,2002(3):23-26.
[15]于金凤,王金信,陈茂学,等. 麦田混生杂草生态经济阈值的研究[J]. 植物保护,2002,28(5):13-15.
[16]Shrestha A,Hanson B D,Fidelibus M W,et al. Growth,phenology,and intraspecific competition between glyphosate-resistant and glyphosate-susceptible horseweeds (Conyzacanadensis) in the San Joaquin Valley of California[J]. Weed Science,2010:58:147-153.
[17]曹春田,李国宇,刘新峰,等. 麦田恶性杂草日本看麦娘的发生及防治[J]. 河南农业科学,2003(10):73.
Research on Growth Rate and Planting Density of Japanese Foxtail (Alopecurusjaponicus) Resistant to Haloxyfop-R-methyl
ZHANG Bin1, LI Jun1,2, DONG Li-yao1,2
(1.College of Plant Protection,Nanjing Agricultural University/Key Laboratory of Monitoring and Management of Crop Diseases and Pest Insects,Ministry of Agriculture,Nanjing 210095,China;2.College of Science,Nanjing Agricultural University/Jiangsu Key Laboratory of Pesticide Science,Nanjing 210095,China)
The germination rate of Japanese foxtail(Alopecurusjaponicus)in the resistant and sensceptible population to haloxyfp-R-methyl could reach to more than 90% after 6 days for water soaking at 4 ℃,but dormancy breaking in the resistant population needed more time than that of sensceptible population.The resistant population presented a lower growths rate than that of sensceptible population after 15 days of sowing,but after that date the growth rate in resistant poputation was getting higher than that of sensceptible poputation. The resistant population began to appear the second leaf after 20 days for sowing that was later than sensceptible population by 2days,and the rate for the second leaf appearance in both populations could reach to 95% affer 25 days of sowing which proved that the resistant population grew more rapidly than that of sensceptible population during the later period of growth time after sowing. The resistant population performed a better growth than that of sensceptible population under the same planting density. Japanese foxtail could grow well under the planting density of 150~200 plants/0.11m2. The suitable growth density for resistant population cowld he designed as 208 plants/0.11m2,but 186 plants/0.11m2for sensceptible population.
Japanese foxtail(Alopecurusjaponicus);dormancy breaking; growth rate; growth densitity
2011-04-11
国家自然科学基金(编号:30971928);国家博士点基金新教师类项目(编号:20090097120046)。
张 彬(1985—),男,硕士研究生,研究方向为除草剂毒理及抗药性。E-mail:2008102117@njau.edu.cn。
董立尧,博士,教授,从事除草剂毒理及抗药性研究。Tel:(025)84395672; E-mail:dly@njau.edu.cn。
S451
A
1003-935X(2011)02-0043-04
猜你喜欢
草业科学(2022年10期)2022-11-18 03:44:18
风流一代·青春(2021年7期)2021-08-09 00:51:21
空间科学学报(2021年1期)2021-05-22 01:36:34
防护林科技(2020年11期)2020-12-30 03:55:36
中国食品学报(2019年3期)2019-01-13 01:47:39
环境保护与循环经济(2017年5期)2018-01-22 02:56:44
散文选刊·下半月(2017年3期)2017-11-07 23:18:28
蔬菜(2016年10期)2016-03-27 12:35:11
中国果菜(2016年9期)2016-03-01 01:28:44
中国蔬菜(2015年9期)2015-12-21 13:04:40
