
猪的分子育种及应用前景
2011-08-06 01:23柳小春
猪业科学 2011年7期
柳小春
(湖南农业大学动物科技学院,长沙 410128)
1 猪种选育方法回顾
世界各国猪的选种方法大体上可归纳为表型选择、基因型选择和标记辅助选择三大类型。
1.1 表型选择
这个阶段经历了很长时间,1960年前基本上都采用表型选择。从遗传学诞生算起,花了60年时间;从我国最早的“伯乐相马”算起,历时2600多年,由于使用方便,不需要特别的仪器和计算,现在仍被广泛使用,特别是外形选择,仍是客户在选购种猪时的一种主要选种方法。应该说,外形在某种程度上可以反映生产性能高低,如前面所说的父系和母系猪的不同外形要求。就反映了父系与母系猪不同的生产性能要求。因此有必要对不同品种、不同用途的种猪提出较可操作的外形标准,而且要着重与生产性能关系大的外形性状。而不过多地拘泥于“猪毛蒜皮”。还需指出,外形选择毕竟是一种“见好就留”的表型选择,没有涉及遗传基础,因此遗传进展较慢。
1.2 基因型选择
随着对基因型有关理论的了解和计算机技术的发展,人们可以透过复杂的表面现象(表型与表型值),剖析其遗传基础,估算其能遗传的部分。为了取得准确可靠的数据,性能测定制度应运而生,包括生长发育性能测定、繁殖性能测定和肥育与肉质测定。与此相应,研制了一批新的电子设备,如测活体背膘厚的超声波测膘仪,测背膘及眼肌面积的B超,测料重比的种猪性能测定自动化系统。法国ACEMO公司推出的ACEMA64新一代自动化性能测定系统,可以准确记录自由采食条件下群饲猪的个体采食量,料槽内的电子测量系统可使猪的采食量精确到2 g。美国OSBOME公司生产的FIRE系统具有相似的功能,除了可测采食时间、采食量外,还可测猪的体重。并通过计算机自动计算,形成猪测定期的日增重、采食量、饲料转化率等数据报表。这套设备虽然投资较大,但可以解决一直困扰人们的群饲猪的料重比指标,还可分析采食行为和计算日增重,为选择提供更准确有效的数据。大量测定数据仍然是表型值,但可通过BLUP法估算出育种值,还可利用遗传评估体系进行场内个体间遗传评估和场问联合遗传评估。场间联合遗传评估必须以准确的数据信息和遗传联结为前提,这方面的工作虽然在我国已试点多年,但困难较多。
1.3 标记辅助选择
1986年后,随着分子生物技术的发展,人们逐渐将DNA重组技术、转基因技术、分子标记技术用于猪育种领域,形成了分子育种技术。将有利基因的分子标记用于选种,形成了标记辅助选择。
2 分子育种技术主要领域
2.1 标记辅助选择
标记辅助选择(marker assisted selection, MAS)是目前应用最多的一个领域,将现代分子生物技术与常规育种方法相结合,借助分子标记选择某一座位基因来改变该座位基因频率的过程。其主要优点是可以用于肉质性状和抗病力等活体很难度量或活体测量花费很大的性状,以及繁殖等在生命活动中表达较晚的限性性状。由于MAS不易受环境影响,且不受性别、年龄限制,可以进行早期选择,从而缩短世代间隔,提高选择强度,提高选种效率和准确性,因此在动物育种中具有广阔的应用价值。
2.2 标记辅助渗入
标记辅助渗入,或称标记辅助导入(marker assisted introgression, MAI)。就是通过遗传标记将某一基因从一个品种渗入到另一个品种,这不同于改良品种某一缺陷时采用传统的导入杂交方法。在MAI时,当供体品种与受体品种杂交后,在每次回交前先根据标记信息选出携带有渗入基因的个体,并用这些个体与受体品种进行回交,可以保证在回交过程中要渗入的基因不会丢失,大大提高基因渗入的效率。Hanset等(1995)已经将大白猪的正常的等位基因固定在皮特兰中,并在3次回交后获得氟烷阴性的皮特兰品系。
2.3 标记辅助保种
畜禽遗传资源保护是世界各国面临的艰巨任务。据FAO(1995)统计,全世界有猪品种353个,其中亚太地区占157个。我国猪的品种数约占亚太地区的一半。目前,全球处于濒危状态的品种约占26%。保种的方法不外乎活体保种(保全套基因)或所谓系统保种(保有利基因)。由于保种经费的拮据,使保种群不可能太大,这就大大增加了基因丢失的可能性。利用整个基因组标记进行监控,可在保种中使基因丢失的概率大大减少,这就是标记辅助保种。这对于防止基因丢失,特别是一些濒危品种的遗传特性的保护有重大现实意义。
2.4 标记辅助抗病育种
疾病不仅给畜牧生产带来巨大经济损失,也危害生产性状的遗传改进,使选择的效率降低。疾病的防治虽可采用免疫接种、药物、卫生和隔离等措施,但从遗传上根治某些疾病,始终是人们追求的目标。目前已发现猪的疾病主基因或遗传标记有位于 6q1.1-1.2 的猪应激综合征的RYRI基因,位于13q3.1区段的E.coliF4的受体基因,位于6q1.1区段的与E.coliF18受体位点紧密连锁的α(1,2)岩藻糖转移酶基因FUT1、FUT2,以及与疾病抗性有关的位于7号染色体上的SLA基因等。利用与疾病抗性有关的基因或基因标记进行抗病力选择,实行标记辅助抗病育种,将可大大加快抗病育种进度,达到根除某些疾病的目的。
3 分子育种的应用前景
3.1 通过标记辅助选择提高猪的产仔数
为了比较不同基因对猪产仔数效应大小,在相同的大白 (158头)、长白 (224头)猪群中采用PCR-RFLP法进行了ESR、FSHβ、PRL、PRLR、NCOA15种与产仔数相关基因的基因型频率检测及不同基因型的总产仔数和产活仔数效应分析,结果表明,对相同母猪群产仔数影响效应最大的是PRLR和NCOA1基因,AA型比BB型母猪总产仔数高2.28~3.33头(P<0.01),产活仔数高1.57~3.30头(P<0.01),其次为ESR和FSHβ基因,BB型比AA型母猪总产仔数高0.55~1.18头(P<0.05,长白例外),产活仔数高0.37~1.20头(P<0.05)。PRL基因对产仔数效应不显著(见表1)。
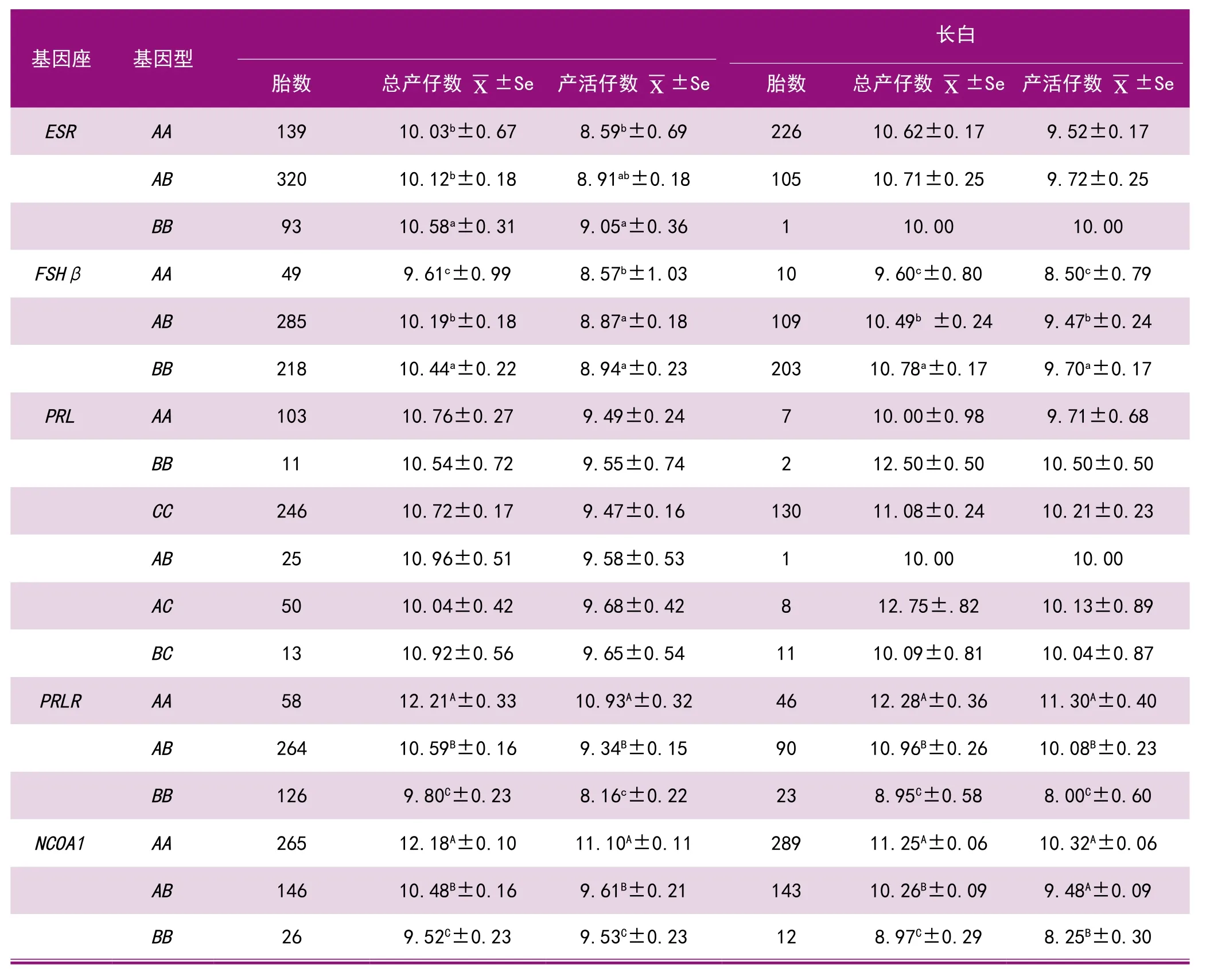
表1 5种基因不同基因型的总产仔数与产活仔数
猪的产仔数是一个非常重要的经济性状,但由于该性状是遗传力低的限性性状,常规选择很难奏效。采用分子标记辅助选择,则可大大加快产仔数的提高。世界著名动物遗传育种学家Chris Haley估计,猪的产仔数提高1头,可给英国养猪业每年增加7亿英磅利润,而整个欧盟每年至少可增利20亿英磅,我国现有约5000万头母猪,采用此项技术使每头母猪每胎增加1头,则每年可增加约800亿元人民币利润,若以推广采用面10%计,每年也可增收80亿元。
3.2 通过标记辅助选择提高猪的生长速度
杜洛克猪在杂交体系中是著名的父系品种,主选性状为生长速度和背膘厚。为此,我们在杜洛克父系选育中检测了PIT1、HDAC1、UCP3、IGF2、MC4R基因的多态性,估算了这些基因的不同基因型对生长性状的效应(见表2)。
由表2可见,经显著性检验,PIT1基因CC型达100 kg活重日龄最小(176.29 d),活体背膘最薄(9.92 mm),生长性能指数最高(108.74),CC型为有利基因型。HDAC1基因GG型达100 kg活重日龄最小(175.41 d),校正活体背膘厚最薄(13.10 mm),生长性能指数最高(104.01),GG为有利基因型。UCP3基因EE型的校正活体背膘厚最薄(12.16 mm),生长性能指数最高(104.55),EE为有利基因型。在IGF2基因座,AA型达100 kg活重日龄最小(173.98 d),BB型的活体背膘厚最薄(15.79 mm),AA型的生长性能指数最高(102.10),综合评价,AA为有利基因型。在MC4R基因座,AA型达100kg活重日龄最小(167.71 d),生长性能指数最高(106.28),AA为有利基因型。
3.3 剔除氟烷敏感基因防治猪应激综合征
氟烷敏感基因是猪中发现最早的主基因之一,该基因的隐性纯合子(HalnHaln)易产生应激综合征(PSS),Haln还有导致产仔数下降、生长慢、胴体短、瘦肉率高和发生PSE肉的效应,给世界各国造成很大经济损失,美国由于PSE肉造成的经济损失每年在3.2亿美元以上,分子生物技术研究表明,PSS是一种分子缺陷病,其机制是位于第6号染色体上q1.1~1.2的CRC(或称RYRI)cDNA上的第1843碱基由C突变成T,造成编码氨基酸由Arg615变成Cys615,根据这一改变目前已可设计一对引物,用PCR-RFLP法得到3种不同基因型图谱,为养猪业中及早检出氟烷敏感基因携带者(HalNHaln)和纯合子(HalnHaln)带来极大方便。我省在引进的杜洛克猪群中发现Haln基因频率达0.3766,正在通过检测剔除。
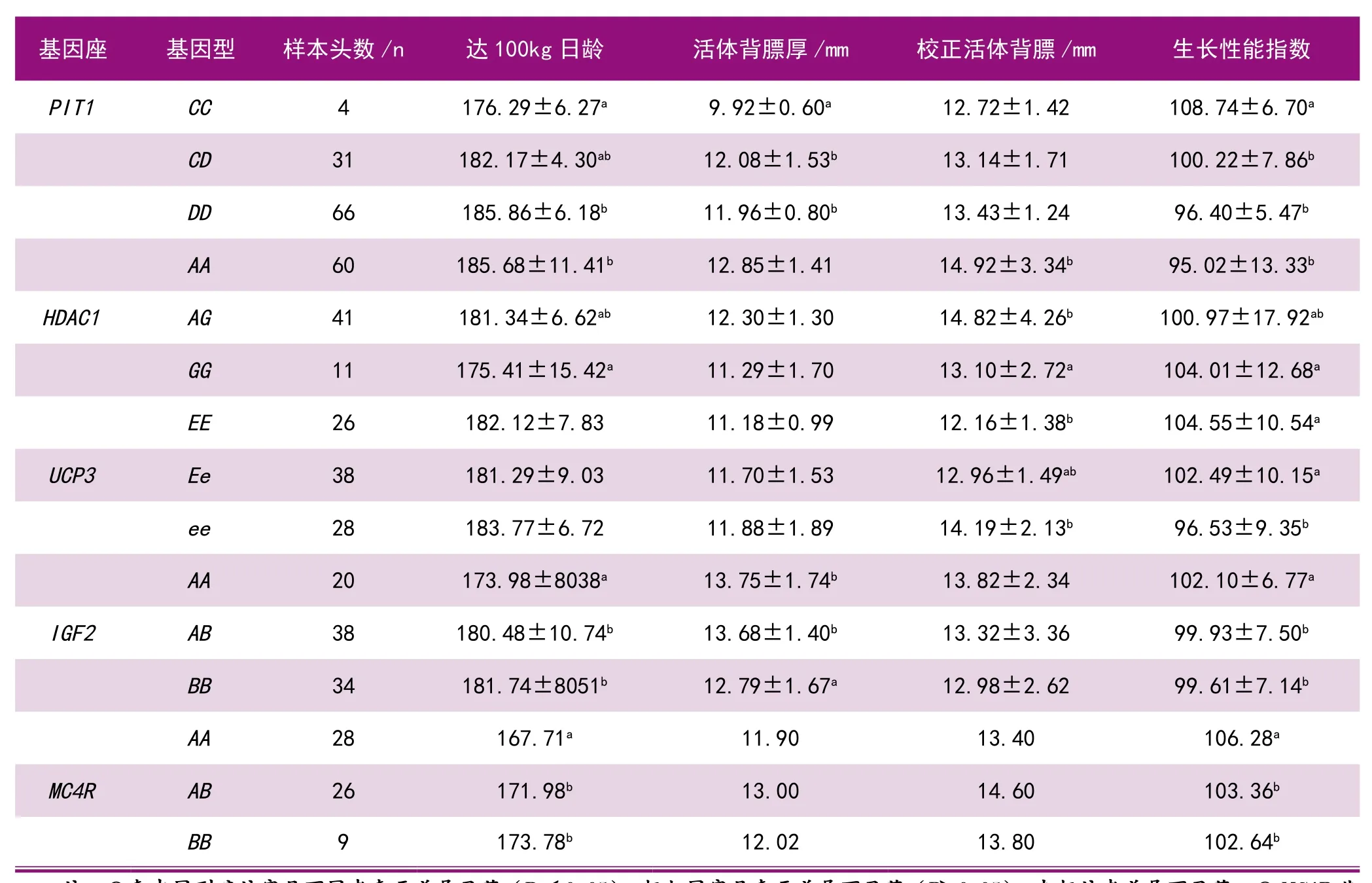
表2 5种基因对杜洛克猪生长性状的效应估计
3.4 肠毒素大肠杆菌病的抗病育种
猪病给养猪生产带来的经济损失约占总产值的12%~15%,在猪病中,仔猪泻痢是最普遍而重要的疾病,由腹泻使仔猪的死亡率达11%~29.5%,其中由肠毒素大肠杆菌(enterotoxigenieE.coli,ETEC)引起的发病率和死亡率约占56.2%和24.7%。值得提出的是ETEC的抗原特性是可变的,已发现的E.coli菌体抗原血清型就有100多种,给研究疫苗及药物治疗带来很大困难。因此,从根本上开展对ETEC抗性遗传机制研究,进行抗病育种已成为近几年的一个研究方向。ETEC有很多种抗原型,其中危害最大的主要有引起1~2周龄仔猪黄、白痢的E.coliK88(F4)和引起断奶后仔猪腹泻和水肿病的E.coliF18。已知E.coliF4是否致病决定于仔猪小肠黏膜上皮细胞有无受体,无受体的猪在受到E.coliF4感染时不发病,表现为抗性;有受体的猪则表现发病。而有无受体受一对等位基因控制,显性基因S控制有受体,隐性基因s为无受体。据此机制,寻找控制受体的基因或标记已成为解决这个难题的关键。为此,我们已从多方面开展了研究,取得了初步成效。
ETEC F18具有类似的遗传机制,目前进展更快,已找到α(1)岩藻糖转移酶基因(FUT1)可作为ETEC F18受体的候选基因。该基因的Hin6I-RFLP位点的M307处G突变为A时,形成抗性型AA,而GG、AG为敏感型。我们检测中外猪种867头样品发现,总的来说抗性型个体(AA)较少,在大白、长白、杜洛克猪中平均为6%,其余全部是敏感型(GG、AG),而本地猪种宁乡猪、大围子猪、沙子岭猪全部属敏感型,无抗性型。这一结果与这些猪种均易感染ETEC病的实际情况一致。但证明了ETEC是可以通过遗传学方法控制的,只要及早对猪只采样检测分型,将AA型留种,淘汰或逐步淘汰GG型、AG型,久而久之,将可使AA型频率升高,GG、AG型频率下降,达到抗病育种之目的。
(略)
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
四川蚕业(2022年2期)2022-11-19
中国动物保健(2021年10期)2021-11-23
中国畜牧杂志(2020年8期)2020-08-16
作文·初中版(2019年8期)2019-09-09
农家致富顾问·下半月(2018年1期)2018-10-21
猪业科学(2017年3期)2017-04-06
猪业科学(2016年4期)2016-01-31
中国医疗美容(2015年4期)2015-04-27
中国医疗美容(2015年4期)2015-04-27
