
岷江上游干旱河谷区岷江柏对不同海拔的响应
2011-06-26 02:08施广
四川林业科技 2011年6期
施 广
(四川省巴州区林业局,四川 巴中 636001)
岷江柏(Cupressus chengiana)是我国珍稀濒危、单型属孑遗种和川甘区域的特有种。岷江柏对土壤pH值的要求范围是中性至碱性,抗旱能力强。其根系发达,能深入岩缝中,固定能力强,所以坡度对岷江柏的生长影响不大,其从平地到悬崖上均可生长。岷江上游干旱河谷区是岷江柏主要分布区之一,该区是长江上游重要的集水区和水源涵养地之一,同时也是我国生物多样性保护的关键地区之一。但该区岩层破碎,地表风化较强,现代地貌过程十分活跃,光、热、水在垂直方向上发生明显的变化,垂直气候显著,由于焚风效应显著,蒸发量远远大于降水量,土壤水分的变化是限制长期定居植物种类、植被类型和植物数量,同时也是控制植被恢复成效的主要因素[1]。严重制约植物生长,从而影响植被的恢复与重建[2]。特别是随着人口的增加和社会经济的发展,干旱河谷生态系统承载了更加严重甚至是前所未有的频繁干扰和过度利用,成为岷江上游山地生态系统退化最严重的生态类型。因此,开展海拔高度变化对岷江柏生理响应的研究,有助于探索岷江柏在不同海拔高度的响应和适应机制,为提高岷江柏造林成效及在造林工作中应采取的技术措施提供参考依据。
1 试验地概况
试验区设在四川省西部理县干旱河谷的典型地带杂谷脑河支流的甘堡乡熊耳村熊耳山,海拔2 000 m~2 500 m,是典型的岷江上游干旱河谷阳坡区域,属于典型的高山峡谷区,地质结构属龙门山断裂带中段。该区土壤以旱生灌木草丛植被下发育的山地燥褐土为主,结构紧实致密,故通气透水和蓄水肥性较差。该地区最高极温37℃左右,最低极温-19℃左右,年平均气温12℃左右。年平均降雨量584 mm,年平均蒸发量1 399 mm,年干燥度1.6~2.5。当地冬季内陆气候盛行,干燥多风,降水稀少,雨季常受西藏高压和太平洋高压影响,间歇性干旱频繁,尤以伏旱持续时间最长,连续高温一般20余d,多者达 46 d。
2 材料与方法
2.1 试验材料
3 a生岷江柏幼苗。
2.2 试验内容与方法
(1)野外采样
实验区海拔设在2 000 m~2 500 m处(理县岷江柏人工造林主要海拔分布范围),布设3个采样点,每个采样地点海拔相差100 m左右。样品采集的布点方法采用随机布点方法。在每个采样小区内的采样点上,采集5处~10处的植株混合组成一个代表样品。采好的样品作好标记,装入液氮罐,带回实验室进行分析。
(2)室内分析
通过对生长在干旱河谷地区不同海拔高度下岷江柏的SOD、POD、脯氨酸、可溶性糖、丙二醛、叶绿素、可溶性蛋白等指标测量,得出不同海拔高度对干旱河谷区岷江柏生长的影响。
游离脯氨酸的测定,采用酸性水合茚三酮法;可溶性糖的测定,蒽酮比色法;丙二醛含量的测定,分光光度法;叶绿素的测定,分光光度法;SOD活性的测定,据SOD抑制氮兰四唑(NBT)在光下的还原作用来确定;可溶性蛋白的测定,考马斯亮蓝G-250法。
2.3 数据处理
利用Exce12003对各处理所测定的生长和生理生化数据进行处理及绘制图表,用SPSS17.0软件对各处理所测定的生长和生理生化数据进行方差分析及主成分分析。
3 结果与分析
3.1 不同海拔土壤水分含量的变化
由于岷江上游干旱河谷地形复杂,土壤异质性强,局部地区气候特殊,随海拔高度的增加,土壤、气候等因子的变化较大。随着海拔高度的增加,气候因子变化有一定规律性,如降雨量增加,蒸发量减少和气温降低。但土壤性质和结构变化并无规律性,可能土壤质地是决定该区干旱河谷土壤含水量的主要因素[1]。
分析结果表明,不同海拔土壤含水量旱雨季变化趋势一致,各海拔0~30 cm土层的平均含水量均随着海拔高度的升高而增加,雨季明显大于旱季(图1)。因为土壤含水量的变化与大气降水紧密相关[2,3],干旱河谷区降水主要发生在雨季内,这期间大气降水几乎是土壤水分的惟一来源,因而,降水过程在很大程度上决定了土壤水分含量。土壤水分由于受降水及降水再分配、根系分布深度、土壤孔隙度、气候条件等的影响,土壤含水量在垂直空间上表现为一定的动态特征[4]。从不同土层看,0~30 cm土层土壤含水量从上到下的变化趋势是随土壤剖面深度的增加呈不同程度的上升趋势(图1)。这与何其华等对岷江上游干旱河谷研究结果相同。何其华认为,土壤水分随土壤剖面深度的增加呈增加趋势[5]。Singhl等在美国西部科罗拉多州对30 cm、45 cm、60 cm、75 cm和90 cm土层的研究也有相似结论[6]。
3.2 不同海拔岷江柏的光合生理日变化

图1 不同海拔土壤含水量变化Fig.1 Seasonal variation of soil moisture at different altitudes
植物的光合作用易受其生长环境的影响,不同生长环境下的植物其光合特性因生态条件而异。在高海拔环境中,影响植物光合特性的环境因子是多方面的,在不同海拔地带又有许多差异,从而造成植物光合适应性的差异。
3.2.1 不同海拔岷江柏的光合速率日变化
由于各海拔梯度下的环境因子不同,从而各海拔下岷江柏的光合速率也随之变化。由图2可以看出,岷江柏的光合速率在各海拔均呈双峰曲线,上午10:00达到最高。随海拔的增加呈先减小然后逐渐增大的趋势。总体上看,上午光合速率高于下午。

图2 不同海拔岷江柏光合速率的日变化Fig.2 Diurnal variation of photosynthetic efficiency in Cupressus chengiana at different altitudes
上午10:00时净岷江柏光合速率达到最高(图2),然后下降,在午后14:00有所上升,之后一直下降。其中,在海拔2 400 m处岷江柏的光合速率最高,2 100处次之,而2 210 m和2 310 m处变化不大。这与栗忠飞等的研究结果不同,栗忠飞认为,净光合速率及生物量均随海拔的增加而下降[6],这可能与研究区的环境因子有关。随着海拔的升高,植物的生长季会缩短,其光合能力也下降,且同一种植物生长在低海拔和高海拔时,光合作用的差异较显著,往往在高海拔植物光合作用的最适温度都要比生长在低海拔处低[7,8]。
3.2.2 不同海拔岷江柏的蒸腾速率日变化
蒸腾速率指植物在一定时间内单位叶面积蒸腾的水量,它是反映植物水分状况最重要的生理指标。一般植物99%的水分都要蒸发,气孔蒸腾是中生和旱生植物蒸腾作用主要的方式,因此,气孔与蒸腾密切相关。蒸腾速率(Tr)是植物水分状况最重要的生理指标,蒸腾作用在降低叶温中起着重要作用,植物通过蒸腾作用扩散水分以降低叶片温度是对高光强高温的一种适应,以减轻高温环境对叶片造成灼伤。
由图3知,不同海拔岷江柏的蒸腾速率日变化呈现双峰型曲线。在中午12:00达到最低,而在下午14:00是为1天中的最大值。在整个日变化过程中,2 210 m处岷江柏的蒸腾速率午后一直保持在较高的水平,且明显高于其他3个海拔。随海拔的升高,岷江柏的蒸腾速率表现为先升高再下降的趋势。

图3 不同海拔岷江柏蒸腾速率的日变化Fig.3 Diurnal variation of transpiration rate in Cupressus chengiana at different altitudes
3.2.3 不同海拔岷江柏的水分利用率日变化
海拔对植物水分利用率的影响是由于它对光合和蒸腾的影响不同所致,蒸腾和光合作用都随海拔的变化而变化,当海拔梯度接近作物最适生长温度和水分时,光合速率上升幅度较小,甚至有所下降,但蒸腾仍然上升,从而导致单叶WUE下降[8]。随着海拔梯度的变化,大气CO2浓度也随之变化,而大气CO2浓度也是影响植物WUE的重要因子,环境中CO2浓度的增加导致作物叶片的净光合速率的增加及其蒸腾速率的降低,从而提高了作物的水分利用率[9]。
从图4可知,岷江柏水分利用率在不同海拔下均呈单峰型曲线,在中午12:00时最大,然后逐渐减小,表现为随海拔的升高先降低后增加的趋势。其中以海拔2 440 m处的岷江柏WUE最高,而2 210 m处最小。总体上看,上午水分利用率大于下午。
3.3 岷江柏生化指标随海拔的变化
3.3.1 膜脂过氧化随海拔的变化
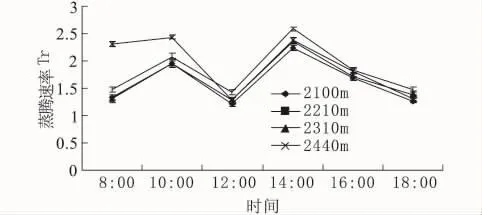
图4 不同海拔岷江柏水分利用率日变化Fig.3 Diurnal variation of water use efficiency in Cupressus chengiana at different altitudes
各种胁迫对植物体的伤害多伴随着植物体内自由基代谢失衡的发生。植物在逆境环境下,其保护酶系统活性增强,以及时清除过多的氧自由基,防止膜脂过氧化,从而维持细胞膜的完整性。也有研究表明,在低温、干旱等环境胁迫下,植物细胞防御系统能力下降,活性氧代谢平衡失调,产生大量的活性氧,从而导致膜质、蛋白质损伤[10]。而MDA作为膜脂过氧化产物,其含量的变化标志着细胞质膜损伤的程度。
从表1可以看出,随海拔的不断升高,岷江柏SOD活性呈先降低后升高的趋势,而POD活性则是随着海拔的升高呈不断下降的变化趋势。这与陈银萍等的研究结果不同。陈银萍等研究表明,随海拔的上升,植物抗氧化酶系统活性呈现增强趋势,随季节变化的低温也使得保护酶系统活性增强[11]。由此可推测,除了水分外,在不同海拔梯度上温度可能也是引起岷江柏抗氧化酶系统变化的主要因素。随海拔的升高,虽然土壤水分含量提高,但是温度却有所下降,因此叶片抗氧化系统在高海拔下的岷江柏随着海拔的升高,植物的生长不仅仅受温度的影响,气温也影响着植物的生长。Ramanjulu等研究发现,高海拔区由于受水分和温度的影响,植物体内活性氧积累超过正常水平,抗氧化酶会由于底物浓度增加而被诱导合成[12]。由此可见,随海拔梯度的上升,酶活性变化是由于植物对恶劣环境适应的结果。
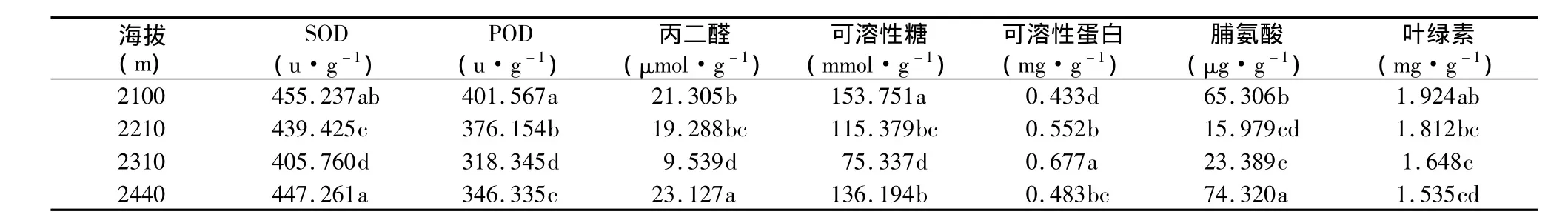
表1 不同海拔生化指标变化Table 1 The change of biochemical indicators at different altitudes
随海拔的不断上升,岷江柏叶片中丙二醛(MDA)含量也呈先降低后升高的趋势,膜脂过氧化在海拔2 100 m处较大,然后随着海拔的升高,在海拔2310m处膜脂过氧化程度达到最低。膜脂过氧化随海拔升高而加剧的原因可能是水分和低温双重胁迫的结果。这说明在干旱河谷区,适合岷江柏生长的地段在海拔2 300 m左右。
3.3.2 渗透调节物质随海拔的变化
植物的抗旱性与渗透调节物质密切相关,渗透调节物质含量的增加,会使细胞原生质的浓度提高,使得植物细胞维持一定的含水量和膨压,从而维持细胞的正常生理功能。
可溶性糖是重要的一种渗透调节物质,对细胞膜也有保护作用,对干旱引起的脱水具有减缓作用。随着海拔的升高,植物大量积累糖类物质和脯氨酸,不但为植株的生长提供了充足的营养成分,而其增加了其细胞的保水能力,进而提高植物的耐逆性。随着海拔的升高,可溶性糖和脯氨酸均呈先下降后升高的变化趋势,而可溶性蛋白则先升高后降低(表1)。在海拔2 100 m处岷江柏的渗透调节物质含量较高,在海拔2 310 m达到最低。这表明在海拔2 300 m左右,岷江柏的水分胁迫较轻,是较适合岷江柏生长的地段。
3.3.3 叶绿素随海拔的变化
叶绿素作为植物光合机构的重要组分,一般来说植物叶片中叶绿素的含量随着海拔的升高而呈现降低的趋势[12,13]。由表1知,随海拔梯度升高,岷江柏叶绿素含量呈下降趋势,但下降的幅度不是很大。这表明高海拔地区的岷江柏虽然生长在恶劣的环境条件下,但其光合作用等生命过程仍能进行,并表现出一定的对不利环境条件的适应性,这可能与其体内的抗氧化系统活性的增强有很大关系。
3.3.4 不同海拔岷江柏的生长指标
一般来讲,低海拔处植物的生长量比高海拔高,这是由于生长季较短和生长速率较低的缘故[13]。从图5可以看出,岷江柏的苗高和地径年生长量均随海拔升高而呈现抛物线变化的趋势,即随着海拔升高生产量先升高,达到一定的海拔后再降低。在海拔2 100 m~2 310 m处,岷江柏苗高和地径年生长量都一直随海拔升高而升高,这除了和各海拔水分相关外,与各海拔的温度可能也有密切的联系。在2 310 m~2 440 m处岷江柏苗高和地径年生长量急剧下降,且直径生长量随海拔升高而降低比植物高的递减要快。

图5 不同海拔岷江柏年生长量变化Fig.5 The annual growth of the height and ground diameter of Cupressus chengiana at different altitudes
有研究表明,岷江柏的适生范围主要在干旱河谷1 800 m~2 500 m左右[14],超过其范围岷江柏的生长就会受到影响,这说明岷江柏的生长除水分条件外,热量条件也是影响其分布的因素之一。由图5知,在海拔2 200 m~2 300 m之间生长最好。因此,在该区营造人工岷江柏林的时候,应该优先选择这一地段。
4 结论与讨论
(1)不同海拔土壤水分含量的变化
不同海拔土壤含水量旱雨季变化趋势一致,各海拔0~30 cm土层的平均含水量均随着海拔高度的升高而增加,雨季明显大于旱季。从不同土层看,0~30 cm土层土壤含水量从上到下的变化趋势是随土壤剖面深度的增加呈不同程度的上升趋势。
(2)不同海拔岷江柏的生长状况
随海拔升高,岷江柏生长发育主要受到温度和水分的限制,因而不同海拔的岷江柏的生长状况有着明显变化。从其生长指标和生理指标来看,在海拔2 200 m~2 300 m之间生长最好。因此,在该区营造人工岷江柏林的时候,应该优先选择这一地段。
(3)研究展望
本研究是在自然干旱胁迫条件下进行的实验,和盆栽实验相比,有较大的优越性,更能客观地反映植物生长的实际情况,但是这样做受季节限制,会耗费更多的人力和物力,所需时间长,工作量大,速度慢,每年结果可比性差,难以重复。另外,由于试验地有雨热同季的气候特征,苗木生长的微环境理化性质会相应的改变,因此植物的生长不仅与水分条件密切相关,还可能与土壤气温及土壤理化性质有关。而本实验只考虑了水分对林木生长的影响,没有考虑气温及土壤理化性质等因子。因此,在研究不同海拔梯度对岷江柏生长的影响时,土壤温度及土壤理化性质等还待进一步研究。
[1] 何其华,何永华,包维楷.岷江上游干旱河谷典型阳坡海拔梯度上土壤水分动态[J].应用与环境生物学报,2004,10(1):68~74.
[2] 王彦辉.在不同植被条件下陇东地区雨季土壤水分消退规律[J].水土保持学报,1989,3(2):85 ~88.
[3] 刘鹄,赵文智,何志斌,等.祁连山浅山区不同植被类型土壤水分时间异质性[J].生态学报,2008,28(5):2389 ~2394.
[4] 张涛,车克钧,王辉.祁连山青海云杉林不同海拔梯度土壤水分动态变化[J].湖北农业科学,2009,48(5):1107 ~1111.
[5] Singhl J S,Milchunas D G,Lauenroth W K.Soil water dynamics and vegetation patterns in a semi-arid grassland[J].Plant Ecology,1998,134:77 ~89.
[6] 栗忠飞,郑 征.西双版纳不同海拔4种雨林幼苗的光合特性和生物量[J].热带亚热带植物学报,2009,17(6):519 ~527.
[7] 朱万泽.贡嘎山地区黄背栎光合作用日变化及光合响应[J].东北林业大学学报,2005,33(6):14 ~18.
[8] 江云,马友华,陈伟,等.作物水分利用率的影响因素及其提高途径探讨[J].中国农学通报,2007,23(9):269 ~273.
[9] 王有科,刘冰,郭凤霞.花椒品种抗冻性比较研究[J].中国沙漠,2006,26(6):1041 ~1045.
[10] 陈银萍,张满效,陈拓.圆柏属常绿木本植物叶片抗氧化系统季节变化与抗冷冻的关系[J].生态学杂志,2006,25(11):1318~1322.
[11] Ramanjulu S,Bartels D.Drought and desiccation-induced modulation of gene expression in lants[J].PlantCell&Environment,2002,25:141 ~150.
[12] 周党卫,朱文琰,滕中华,等.不同海拔珠芽蓼抗氧化系统的研究[J].应用与环境生物学报,2003,9(5):489 ~492.
[13] 潘红丽,李迈和,蔡小虎.海拔梯度上的植物生长与生理生态特性[J].生态环境学报,2009,18(2):722 ~730.
[14] 袁志忠,何丙辉.岷江柏种群现状及研究进展[J].山区开发,2003,6:34~35.
猜你喜欢
散文诗(2021年22期)2022-01-12
散文诗世界(2021年12期)2021-12-17
环球时报(2019-05-23)2019-05-23
岷峨诗稿(2019年4期)2019-04-20
少儿美术(快乐历史地理)(2018年7期)2018-11-16
阿来研究(2018年1期)2018-07-10
高原山地气象研究(2016年2期)2016-11-10
廉政瞭望(2016年13期)2016-08-11
河北遥感(2014年4期)2014-07-10
塔里木大学学报(2014年3期)2014-03-11
