
嘉陵江上游低山暴雨区3种不同植被类型氮循环研究
2011-06-26 02:08吴雪仙龚固堂张发会陈俊华朱志芳慕长龙张启珍
四川林业科技 2011年6期
吴雪仙,龚固堂,张发会,陈俊华,朱志芳,慕长龙* ,张启珍
(1.四川省林业科学研究院,四川 成都 610081;2.理县林业局,四川 理县 623100)
氮是组成蛋白质和核酸的基本元素,是植物生长和发育所需的大量营养元素之一,也是植物主要从土壤中吸收的元素[1]。森林生态系统中的氮素循环是氮素通过不同途径进入森林生态系统,再经过许多相互联系的转化和移动过程后,又不同程度地离开这一系统。这一循环是开放性的,它与大气和水体等外界环境进行着复杂的交换[2]。由于氮素为非矿质元素,土壤中可利用氮容易缺乏,从而导致氮成为许多地区树木生长最重要的限制性因子[3~6]。而有些地区如北美和欧洲,“氮饱和”明显地影响了森林生态系统的正常结构和功能[7~9]。在美国南部,砍伐或火烧导致森林土壤中NO-3从溪流水中大量流失,并引起一系列连锁反应,如土壤酸化,溪流中SO2-4浓度的下降等[10]。近几十年的研究
表明,氮一方面是许多森林生产力和产量的重要限制因素,另一方面又与生态环境的恶化,如酸雨、温室效应、地下水中NO-3的污染以及生物多样性丧失密切相关[11~14],同时还与 C、S、P 等元素的循环具有明显的耦合作用[15]。森林生态系统中土壤氮通常占整个生态系统氮贮量的90%以上;且森林土壤氮转化所提供的占土壤全氮1%~5%左右的无机氮才是植物吸收和造成环境污染的最主要的形式[16,17]。因此,了解氮的循环过程和机制,对正确地解释和调控森林生态系统中氮及其养分循环,科学地经营管理森林生态系统,维持生态系统的健康,都具有十分重要的意义。
本文通过对嘉陵江上游低山暴雨区3种不同植被类型(湿地松纯林、刺槐纯林、湿地松与刺槐的混交林)的氮素的凋落物归还、降雨淋溶归还、地表径流氮素的输出、土壤氮素含量等方面进行研究,旨在了解不同植被类型的氮循环机制及其对氮素的利用效率,为维护地力和提高林分生产力及其可持续发展提供理论依据。
1 研究区概况
研究区设置在广元市城北4 km的工农镇碗厂沟,行政区隶属于广元市中区工农镇,东经105°54'24″~105°58'48″,北纬 32°27'51″~ 32°31'39″,位于长江支流嘉陵江上游,地处四川盆地北缘,米仓山南麓,属于秦巴山余脉低山暴雨区,东西长11.8 km2,南北宽约2.79 km2,地势由东北向西南倾斜,海拔515 m~815 m。
本区受秦岭和大巴山的影响,属于北亚热带湿润季风气候,多年平均气温16.1℃,极端最高气温为41.5℃,极端最低气温为 -3.5℃,≥10℃积温5 065.4℃,无霜期263 d,年平均降雨量达937.2 mm;降雨年际差异大,最高年降雨量1 518.1 mm(1961年),最低年降雨量为580.9 mm(1975年);年内降雨分配不均匀,主要集中在6月~9月,占全年降雨量的72.5%;年平均出现暴雨3.3 d,大暴雨0.6 d,区内多暴雨,雨量占全年降雨量的40%以上。由于降雨集中,常出现冬干、春旱、初夏旱、盛夏暴雨等自然灾害。
土壤主要为砂岩、石灰岩发育形成的黄壤,pH值4.5~6.5,土壤干燥紧结。另外,紫色土也有分布。
现有植被主要为人工林和次生灌草植被,人工林有麻栎林、刺槐林、桤木林、马尾松林、湿地松林,次生灌草植被主要由槲栎、火棘、铁仔等灌木树种和黄茅、白茅、莎草等草本植物组成。
2 研究方法
2.1 标准地的设置
本研究选择具有代表性的植被类型(湿地松林、刺槐林、湿地松×刺槐混交林)作为研究对象,采用典型抽样法设置林分标准地,面积为20 m×20 m。其基本情况见表1。

表1 标准地基本情况Table 1 Basal situation of standard site
2.2 材料收集与处理
2.2.1 土壤样品的采集
森林土壤样品的采集与制备参考GB7830-87。
在样地外选择3处具有代表性的地点挖掘土壤剖面,每隔20 cm取样,从下往上取。每层采集土样1 kg左右,带回实验室风干、研磨、过筛、混合分样,储存供化学分析用。根据每层土壤的重量和含氮量计算氮的储量。
2.2.2 降雨样品收集
在每个标准地内设置穿透雨和树干径流收集器各6个,在附近空旷地上设立6个降雨收集器,将每月的降雨取样带回试验室分析,测定其N元素的含量。
2.2.3 生物量的测定
在各标准地进行每木检尺,根据每木检尺的结果,确定各植被类型的平均胸径和平均树高,选择2株~3株平均标准木,按其平均营养面积将其盘根挖倒,记录胸径、树高。按Monsi分层切割法,以2 m为一区分段直接称取标准木各段树干(带皮)、枝叶果的鲜重,通过在分段处截取圆盘和在树冠所在区分段选取标准枝,求得干与皮、枝与叶比例以推算各组分(器官)鲜重并在各组分中均匀取样,带回室内称量,并在65℃恒温下烘至恒重,求得各组分干湿比换算得各组分生物量(干物质质量,下同)。根系按根颈、粗根(>5 mm)中根(2 mm ~5 mm),细根(<2 mm)分类称量,如前法取样求干湿比,得出各标准木根系的分类生物量。在做生物量的同时采集各器官样品供化学元素分析。
2.2.4 凋落物的收集
凋落物的收集采用收集网法。在各标准地内分别沿对角线的4个角和中心位置设立5个0.5 m×0.5 m的凋落物收集框架,每月定期收集网中的凋落物,以标准地为单位装袋并挂上标签带回实验室测定凋落物量,并分为叶﹑枝﹑树皮﹑花果等组分,分别置于65℃恒温箱下烘干至恒重,然后称重。
2.3 样品分析
2.3.1 植物样品的测定
植物样品用硫酸—高氯酸消煮,然后用凯氏定氮法测定全N,参考GB7886—87至7888—87。
2.3.2 土壤样品的测定
森林土壤样品的测定参考 GB7848—87至7865—87;
全N用定氮蒸馏法。
3 结果与分析
3.1 不同植被类型林下土壤氮储量
不同植被类型林下土壤的全N储量见表2,可见3种林型均是表层土壤的N储量最高,40 cm~60 cm的N储量最低。3种类型林下全N储量的总量是混交林最大,其次是刺槐林,最小的是湿地松林。
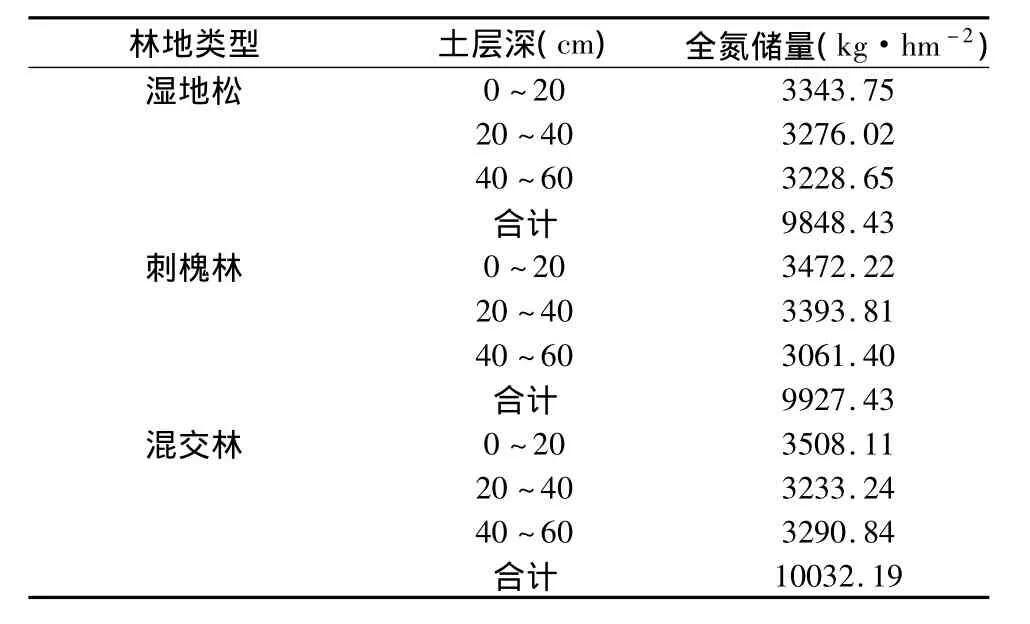
表2 不同植被类型林下土壤的全氮储量Table 2 The nitrogen stock in soil of the different vegetation types
3.1 不同植被类型各组分的氮含量
植物中营养元素的分布特征一方面反映了植物自身的特性,是植物长期演化的结果;另一方面,受所处生境的影响,生境中的土壤肥力状况是影响植物生长发育及元素分布的重要因素[18,19]。由于植物不同器官的生理机能不同,不同营养元素在植物体内的功能不同,营养元素在植物不同器官及不同营养元素在同一器官中的分布也有差异。3种植被类型各组分N含量的变化范围在0.758 g·kg-1~14.98 g·kg-1,3种植被类型各组分N含量的大小不同,其各组分的N含量大小顺序,湿地松林为叶>根>皮>枝>干;刺槐林为叶>果>根>皮>枝>干;混交林的湿地松为叶>根>枝>皮>干;混交林的刺槐为果>叶>皮>根>枝>干(表3)。湿地松纯林与混交林的湿地松相比,混交林的枝、叶、根均大于纯林,其中混交林中湿地松根的N含量是纯林的1.97倍,枝和叶分别是纯林1.67、1.53倍;干和皮的N含量是纯林大于混交林,纯林干、皮的N含量分别是混交林的1.42、2.03倍。混交林中刺槐枝、干、皮、果的N含量均比刺槐纯林高,而叶、根的N含量是纯林高于混交林。不论是纯林还是混交林,各组分N含量均是刺槐大于湿地松。

表3 不同植被类型各组分的N含量Table 3 N Concentration of different vegetation types(g·kg-1)
3.2 不同植被类型各组分氮的积累量及其分布
森林生态系统中,乔木层是最活跃、最重要的亚系统,该系统所进行的初级生产既是能量的固定过程,亦是营养元素的积累过程[20,21,22]。林木营养元素的积累与分配主要决定于林木生物量的积累(现存量)及其各组分中营养元素的含量。各组分养分积累量为植被类型各组分的生物量与其相应养分浓度的乘积。湿地松林、刺槐林、湿地松与刺槐混交林N的总积累量分别为136.43 kg·hm-2、210.40 kg·hm-2、169.66 kg·hm-2(表4),以刺槐林 N 积累量最大,其次是混交林,混交林总积累量是湿地松林的1.24倍。在湿地松林中分布在干47.23 kg·hm-2,占总积累量的 34.62%;干皮 13.62 kg·hm-2,占 9.99%;树枝 4.92 kg·hm-2,占 3.61%;叶52.07 kg·hm-2,占 38.17%;粗根、中根、细根分别占总积累量的12.47%、0.25%、0.89%。叶片中积累的N元素最多,其次是干材,中根最少。在刺槐林中分布在干材、干皮、树枝、叶、果、粗根、中根、细根中的积累量分别为20.87 kg·hm-2、29.05 kg·hm-2、32.87 kg·hm-2、16.60 kg·hm-2、2.30 kg·hm-2、101.77 kg·hm-2、3.95 kg·hm-2、3.32 kg·hm-2,分别占总积累量的 9.92%、13.81%、15.62%、7.89%、1.10%、48.37%、1.17%、1.58%,粗根的积累量最多,其次是枝,果的积累量最小。混交林中干材、干皮、树枝、叶、果、粗根、中根、细根的积累量分别为 55.86 kg·hm-2、19.52 kg·hm-2、20.83 kg·hm-2、34.30 kg·hm-2、1.84 kg·hm-2、34.30 kg·hm-2、1.85 kg·hm-2、1.11 kg·hm-2,分别占总积量的 32.93%、11.51%、12.28%、20.22%、0.11%、20.22%、1.09%、0.66%,混交林中以干材的N积累量最多,其次叶和粗根,最小的是果。
各器官N元素的积累量存在明显的差异,3种植被类型各器官的N积累量的大小顺序,湿地松林为叶>干材>粗根>干皮>树枝>细根>中根;刺槐林为粗根>树枝>干皮>干材>叶>中根>细根>果;混交林为干材>叶>粗根>树枝>干皮>中根>细根>果。植物体内不同组分的营养元素浓度差异主要是由于植物体内各组分生理活性不同引起。但组分营养元素浓度的差异并非是各组分营养元素贮量间差异较大的主要原因,而是不同组分间生物量的差异所致[81]。
3.3 不同植被类型凋落物N年归还量
凋落物中的养分归还于森林土壤,是土壤养分的重要来源。3种植被类型N的年归还量列于表5。湿地松林N的年归还量为31.06 kg·hm-2,在各组分中,由于叶凋落量大,因此归还量也最大,占N年归还量的54.82%,果由于凋落量小,归还量也最小,只占N年归还量的0.33%。刺槐林N的年归还量为38.95 kg·hm-2,各组分中,以杂物归还最大,占总N归还量的63.28%,杂物中由于养分含量高,且凋落量也比较多;叶的归还量占其次,枝的归还最小,只占总归还量的1.66%。混交林N的年归还量为40.04 kg·hm-2,与刺槐林相似,也是杂物归还量最大,占总归还量的42.03%,枝的归还最小,仅占4.95%。3种植被类型N的年归还总量的大小顺序为:混交林>刺槐林>湿地松林,说明营造混交林有利于提高凋落物的养分归还量。凋落物归还的养分多少与凋落物量及凋落物的养分含量有密切的关系。
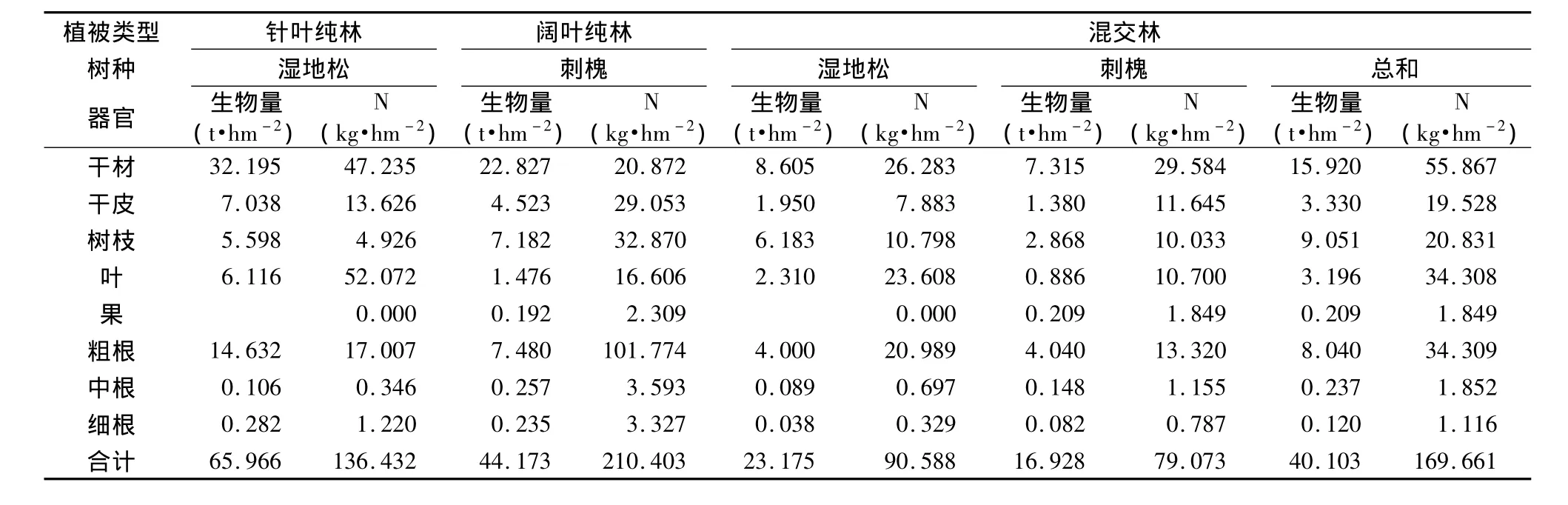
表4 不同植被类型N养分积累量与分布Table 4 N accumulated amount and distribution of different vegetation types
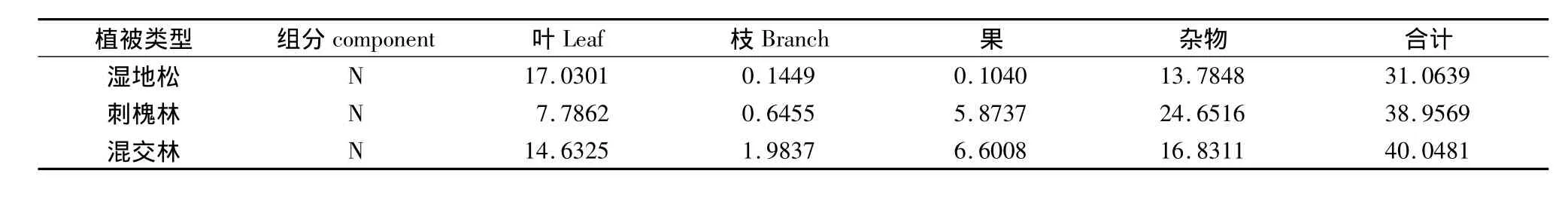
表5 不同植被类型凋落物中N的年归还量Table 5 Annual N return in litter fall of different forests(kg·hm-2·a-1)
3.4 不同植被类型N库特征
从表6可以看出,3种植被类型的总氮量分别为:6 787.27 kg · hm-2,7 115.39 kg· hm-2、6 951.06 kg·hm-2,其中在活体生物量分别为:136.43 kg·hm-2、210.40 kg·hm-2、169.66 kg·hm-2,分别占总量的2.01%、2.96%、2.44%。土壤(0 ~40 cm)中分别为:6 619.78 kg·hm-2、6 866.03 kg·hm-2、6 741.35 kg·hm-2,分别占总量的97.53%、96.50%、96.98%。如果按地上和地下部分划分,湿地松林分别占2.19%和97.81%,刺槐林分别占1.98%和98.02%,混交林分别占2.48%和97.52%。由此可见,3种植被类型的营养元素主要贮存于土壤中。

表6 不同植被类型的氮库特征Table 6 Nitrogen stock of different vegetation types
3.5 不同植被类型降水淋溶氮的输入输出
3.5.1 不同植被类型降水淋溶的氮归还量
降水淋溶是指由于降水作用使得森林植物叶片、树皮以及根系部分的养分被淋洗的过程。在某些气候区,对于某些养分来说,降水淋溶是森林生态系统养分归还的一种主要途径[23]。几乎所有的养分均可从树冠和树干上被降雨淋溶。降雨淋溶的养分归还量大小取决于森林的树种组成和结构。湿地松林、刺槐林、湿地松与刺槐混交林N元素的年淋溶归还量分别为13.08 kg·hm-2,13.10 kg·hm-2,14.14 kg·hm-2(表7),其中混交林年淋溶量归还量最大,湿地松林最小,淋溶归还量的大小与林分的郁闭度和冠层厚度大小有关。从降雨归还的各组分来看,湿地松林、刺槐林、湿地松与刺槐混交林林冠雨的归还量分别为12.649 kg·hm-2、12.550 kg·hm-2、13.291 kg·hm-2,占各林型降水淋溶归还量的96.63%、95.77%、93.99%;湿地松林、刺槐林、湿地松与刺槐混交林树干径流的归还量分别为1.18 kg·hm-2,1.35 kg·hm-2,1.84 kg·hm-2,分别占降水淋溶归还量的3.36%,4.22%,6.01%;表明在降水归还途径中,主要是林冠雨的归还。从雨量级来看,3种植被类型不论是树干径流归还还是林冠归还,均是小雨的归还量最大,暴雨的归还量最小,说明雨量级越大其N归还量越小。

表7 不同植被类型树干径流、林冠水的N归还量Table 7 N return of the runoff and crown of different vegetation forms(kg·hm-2)
3.5.2 不同植被类型降水淋溶氮的输出
随着穿透雨强度的增大和土壤水分饱和,穿透降雨和树干径流进入地被后,养分一方面被枯枝落叶、腐殖质和土壤胶体所吸附固定,另一方面水分又淋溶出枯枝落叶、腐殖质和土壤中的养分,并随地表径流和土壤渗漏水而流失。本研究由于实验条件有限,只考虑了地表径流,没有考虑地下渗流。湿地松林、刺槐林、湿地松与刺槐混交林的地表径流的输出量分别为:0.55 kg·hm-2、0.55 kg·hm-2、1.27 kg·hm-2,湿地松纯林和刺槐纯林N的输出量相差不大,混交林输出量分别是湿地松纯林和刺槐纯林的2.28倍、2.29倍,混交林的养分输出量比较大可能与林下凋落物的现存量较少有关。
3.6 不同植被类型氮元素生物循环特点
生物循环是土壤(基质)与植物间营养元素的流动过程,通过植被的吸收、存留和归还3个不同的生理生态过程来维持平衡。其平衡式为:吸收=存留+归还
生态系统养分循环的特点可以通过吸收系数、利用系数及循环系数来表征.吸收系数是指单位时间、单位面积植物所吸收的某种元素的量与土壤中相应元素总量之比,其大小与土壤中元素的贮存量及植物体对各元素的需求量有关;利用系数为单位时间、单位面积植物所吸收的某种元素的量与存在于植物现存量中相应元素总量之比,反映生态系统元素存贮速率的大小,该系数越大,则表明目前植物对该养分元素的贮存能力越大;循环系数是单位时间、单位面积植物归还量与相应吸收的量之比,反映了元素在循环过程中的残留量大小,循环系数越大,表明元素循环的速率越快,系统中存留量的比例就越小,而流动性越大[24]。
表8反映了不同植被类型N元素的生物循环特征。湿地松林、刺槐林、湿地松与刺槐混交林的年吸收量分别为37.561 kg·hm-2、48.976 kg·hm-2、48.127 kg·hm-2,刺槐林的年吸收量最大,湿地林的吸收量最小,混交林的年吸收量是湿地松纯林的1.28倍。3种植被类型年存留量的大小顺序为刺槐林>混交林>湿地松林;归还量的大小顺序为:混交林>刺槐林>湿地松林,各植被类型的N归还量均大于存留量,说明植物吸收的N元素主要用来构建地上部分。3种植被类型N元素的吸收系数、利用系数、循环系数的大小顺序均是混交林>湿地松林>刺槐林,说明混交林对N元素的贮存能力和循环速率最大。

表8 不同植被类型N元素的生物循环特征Table 8 Biologic cycle characteristics of N element in different vegetation types
3.7 不同植被类型氮元素的生物地球化学循环
3种植被类型的生态系统是一个开放系统,外界的各种干扰因素均会对系统的养分循环产生影响,包括大气输入(干湿沉降入地质和各种生物因素的输出等)[24],由于养分的地质输入在短期内可视为零,在这里仅考虑了大气降水的养分输入和随地表径流的养分输出,由此,计算了不同植被系统养分元素的生物地球化学循环通量(表9)。
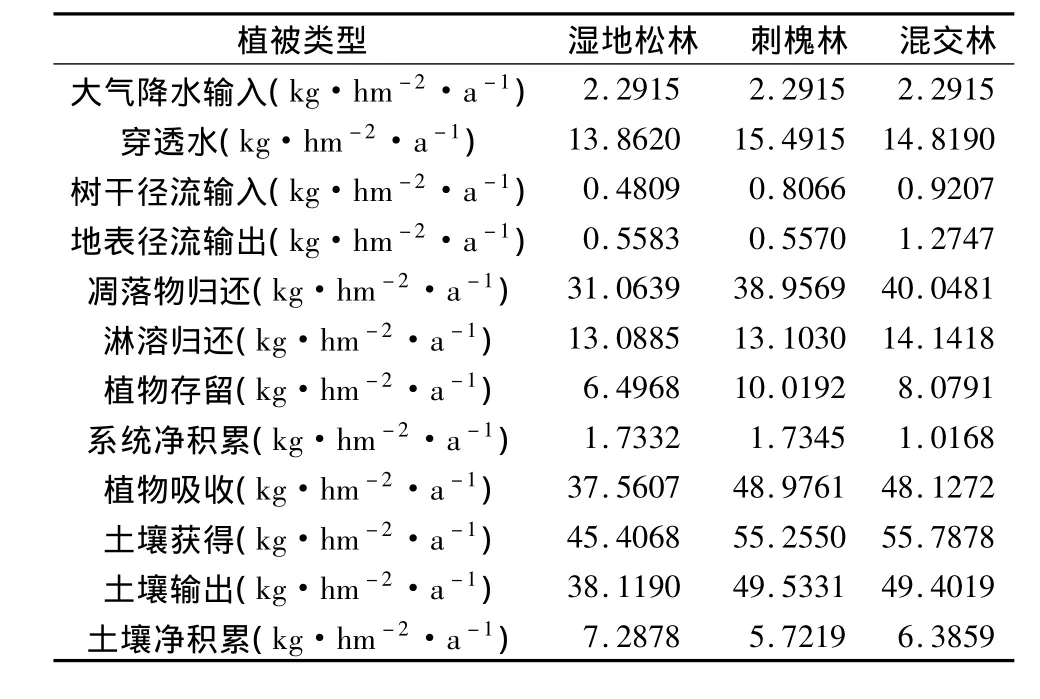
表9 不同植被类型N的生物地球化学循环Table 9 Biogeochemical cycle of N element in different vegetation types(kg·hm-2·a-1)
由表9可以看出,3种植被类型的N输入量均大于输出量,系统处于良好的养分积累状态,主要是由于植被层的良好的覆盖作用,使其具有较好的抗侵蚀作用,使养分流失量较低。土壤中氮的输入量也大于输出量,说明植被不需要外源补给,也能保持持续稳定的发展。3种植被系统N的净积累量的大小顺序为:湿地松林≈刺槐林>混交林,土壤中N的净积累量的大小顺序为:湿地松林>混交林>刺槐林。
4 小结
低山暴雨区3种植被类型均处氮素积累状态的生态系统,系统中95%以上的氮素分布于土壤(0~40 cm)中,生物量中只占2% ~3%。
湿地松林、刺槐林、湿地松与刺槐混交林N元素的年淋溶归还量分别为:13.089 kg·hm-2,13.103 kg·hm-2,14.142 kg·hm-2,混交林年淋溶量归还量最大,湿地松林最小,淋溶归还量的大小与林分的郁闭度和冠层厚度大小有关。在年淋溶归还量中主要是林冠雨的归还,占各植被类型降水淋溶归还量的96.63%、95.77%、93.99%;树干径流的归还量较小,分别占降水淋溶归还量的3.36%,4.22%,6.01%。
生态系统养分循环的特点可以通过吸收系数、利用系数及循环系数来表征,湿地松林、刺槐林、湿地松与刺槐混交林N元素的年吸收量分别为37.561 kg·hm-2、48.976 kg·hm-2、48.127 kg·hm-2,混交林的年吸收量是湿地松纯林的1.28倍。3种植被类型年存留量的大小顺序为:刺槐林>混交林>湿地松林;归还量的大小顺序为:混交林>刺槐林>湿地松林,各植被类型的N归还量均大于存留量,说明植物吸收的N元素主要用来构建地上部分。3种植被类型N元素的吸收系数、利用系数、循环系数的大小顺序均是混交林>湿地松林>刺槐林,说明混交林对N元素的贮存能力和循环速率最大。
3种植被类型的N输入量均大于输出量,系统处于良好的养分积累状态,土壤中养分的输入量也大于输出量,说明植被不需要外源补给,也能保持持续稳定的发展。3种植被系统净积累量的大小顺序为:湿地松林、刺槐林>混交林,土壤中N净积累量的大小顺序为:湿地松林>混交林>刺槐林。
[1] Ingestad T.Plant Growth in Relationto Nitrogen Supply.In:Clark,F.E.&Rosswall,T.eds.Terrestrial Nitrogen Cyles.Ecol.Bull,Stockholm,1981,33(303):268 ~271.
[2] 朱兆良.我国农业生态系统中氮素的循环和平衡[A].见:朱兆良,文启孝主编.中国土壤氮素[M].南京:江苏科学技术出版社,1992:288~303.
[3] 韩兴国,李凌浩,黄建辉.生物地球化学概论[M].北京:高等教育出版社!施普林格出版社,1999,197~244.
[4] 周才平,欧阳华,裴志永,等.中国森林生态系统的土壤净氮矿化研究(英文)[J].植物生态学报,2003,27(2):170 ~176.
[5] Deluca T H,Nilsson M C,Zackrisson O.Nitrogen mineralization and phenol accumulation along afirechrono sequence in northern Sweden[J].Oecologia,2002,133:206 ~ 214.
[6] Wright R F,RasmussenL.Introduction to the NITREX and EXMAN projects[J].For.Ecol.Man,1995,101:1 ~7.
[7] Tilman D.Plant dominance along an experimental nutentgradient[J].Ecology,1984,65:1445 ~1453.
[8] Aber J D,Nadelhoffer K J,Steudler P,et al.Nitrogen saturation in northern forest ecosystems[J].BioScience,1989,39:378 ~386.
[9] Vitousek P M.Gosz J R,GRIERCC,et al.A comparative analysis of potential nitrification and nitrate mobility in forest ecosyststems[J].Ecol.Monogr,1982,52:155 ~177.
[10] Ettershan K G J,Ettershank M B,Whitford W G.Effects of nitrogen fertilization on primary prodctivety in a Chihuahuan Desert ecosystem[J].J.Aridenviron,1978,1:135 ~139.
[11] Fisher R F,Binkley D.Ecology and Management of ForesSoils[M].NewYork:JohnWiley&Sons,2000.
[12] Schlesinger W H.Biogeochemistry:An Analysis of Globaal Change[M].California:Academic Press,1991.
[13] Vitousek P M,Aber J D,HowarthRW,et al.Human alteration of the global nitrogen cycle:causes and consequences[J].Ecol.Appl,1997,7:737 ~ 750.
[14] Binkley D,Hart S C.The components of nitrogen availability assessment in forests soils[J].Adv.Soil Sci,1989,10:57 ~111.
[15] Vestgarden L S,KJnaas O J.Potential nitrogen trans for mations in mineral siols of two coniferous fotests exposed todifferent N inputs[J]For.Ecol.Man,2003,174:191 ~ 202.
[16] Peng S L.Restoration ecology and recovery of degraded ecosystem.Journal of Chinese Academy Sciences,2000,(3):188 ~192.
[17] Zhang X B,Guo X Q,Zhou T L,et al.Floristic analysis of seed plants in Ziwuling forest region.Acta Botanica Boreali-OccidentaliaSinica,2004,24(2):267 ~274,18,19,22 ~24.
[18] Nie D P.Biological cycling of the nutrient element in forest ecosystems.Forest Rresearch,1991,4(4):435 ~440.
[19] Liu G Q,Tu X N,Zhao S D,et al.Distributional characteristics on biomass and nutrient elements of Pine-Quercus liaotungensisforest in MT.Q inling.Scientia Silvae Sinica,2001,37(1):28 ~36.
[20] Huang J H,Chen L Z,Han X G.Changes of nutrient concentration during the decomposition process ofQuercus liaotungensistwigs.Acta Phytoecologica Sinica,1998,22(5):398 ~402,22 ~24.
[21] He C Q,Zhao K Y.The accumulation,allocation and biological of the nutrient elements inCarez lasiocarpawetland.Acta Ecologica Sinica,2001,21(12):2075 ~2080.
[22] 樊后保,苏兵强.杉木人工林生态系统的生物地球化学循环Ⅰ:养分归还动态.应用与环境生物学报,2000,6(2):127~132.
[23] 李绍才,孙海龙.等.岩石边坡基质一植被系统的养分循环.北京农业大学报,2006,28(2):85~90.
猜你喜欢
Plasma Science and Technology(2022年9期)2022-08-29
农业与技术(2021年6期)2021-12-12
现代园艺(2020年2期)2020-12-22
林业与生态(2020年2期)2020-04-16
小溪流(故事作文)(2018年10期)2018-12-28
现代园艺(2018年2期)2018-03-15
连环画报(2016年10期)2016-12-16
心潮诗词评论(2016年1期)2016-12-01
山东林业科技(2016年5期)2016-07-05
乡村地理(2016年2期)2016-06-15
