
结球甘蓝抗寒性配合力分析及优良抗寒组合选育
2011-05-22 03:55张扬勇方智远刘玉梅杨丽梅孙培田
中国蔬菜 2011年14期
张扬勇 靳 哲 方智远 刘玉梅 杨丽梅 庄 木 孙培田
(中国农业科学院蔬菜花卉研究所,北京 100081)
结球甘蓝(Brassica oleracea var.capitata L.)简称甘蓝,为十字花科芸薹属蔬菜,在蔬菜的周年供应及出口贸易中起重要作用,近年来全国种植面积约90万hm2(杨丽梅 等,2011)。随着市场需求的变化,甘蓝产销发生了一些新变化(方智远,2008)。其中一个新变化就是长江中下游地区的越冬甘蓝得到迅速发展,仅湖北省嘉鱼县越冬甘蓝种植面积就达 1万 hm2左右。该茬口一般在7~8月播种,翌年1~3月收获,因此要求甘蓝品种可耐-5 ℃左右的低温,抗病耐裂性强。鉴于该茬口品种主要依赖于进口种子,中国农业科学院蔬菜花卉研究所于21世纪初启动了抗寒甘蓝新品种选育工作,目前已经选育出了一批优良抗寒自交系,这些自交系间抗寒性存在一定的差异,但是他们的抗寒性配合力还不清楚。
关于抗寒性配合力的研究,已经在小麦(孙贵臣和徐环澄,1993)、水稻(Cruz et al.,2006;Cruz et al.,2010)等作物上报道,但尚未见关于甘蓝抗寒性配合力的报道。在甘蓝中已有耐裂球性、产量、抗病性等性状的配合力分析(Chiang,1972;方智远和孙培田,1982;More & Wallace,1988;庄木 等,2009;李梅 等,2009),这些性状的配合力测定为常规育种提供了科学依据。本试验以本所选育的6个甘蓝抗寒自交系为材料,采用Griffing双列杂交方法(Ⅳ)测定甘蓝抗寒性的配合力并进行分析,为甘蓝抗寒新组合的选育提供依据。
1 材料与方法
2010年春季选用6份甘蓝抗寒自交系09-372、09-398、09-386、09-344、09-347、09-362(分别编号为A、B、C、D、E、F),按照Griffing双列杂交方法(Ⅳ)配制15个杂交组合。2010年秋季在本所顺义试验农场露地种植,采用随机区组设计,3次重复,小区面积3.5 m2,每小区定植13株,7月16日播种,8月20日定植。
自然降温至最低温度0 ℃以下时,开始进行抗寒性调查,分2次进行:11月10日(寒害前期)、11月22日(寒害后期)。每小区调查全部植株的寒害级别,并计算寒害指数。寒害分级参照李锡香和方智远(2007)结球甘蓝幼苗的寒害分级标准,并结合甘蓝植株的田间植物学性状制定了寒害级别:0级,无任何受冻症状;1级,植株外部 1~2片叶受冻;3级,植株外部 3~5片叶受冻;5级,植株外部6~8片叶受冻;7级,植株全部叶片受冻萎蔫;9级,植株全部外叶枯死。
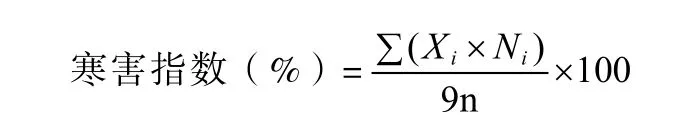
式中,Xi表示寒害级别;Ni表示受低温伤害后相应级别的植株数;n为小区总植株数。
根据 Griffing提出的双列杂交配合力统计原理,按照刘来福等(1984)的计算方法,利用DPS软件对小区寒害指数进行统计分析。
2 结果与分析
2.1 抗寒材料的选育
近10年来,本所甘蓝课题组先后引进寒春4号、M-3、帕特等甘蓝优良抗寒一代杂种逾50份。其中部分品种为胞质不育系配制而成,无法加以利用,因此笔者选择了以自交系配制的优良抗寒品种进行自交分离和田间抗寒性筛选,已选育出了抗寒自交系逾80份,其中球形、球色、球内结构、抗病性等经济性状优良的自交系逾30份,选取其中6份抗寒性较好但又有一定差异的优良自交系进行配合力测定(表1)。

表1 6份甘蓝抗寒自交系的主要农艺性状
2.2 杂交组合的抗寒性表现
从表2可以看出,A×B、B×E、D×E、C×D等4个组合在两次调查中的平均寒害指数均较低,抗寒性好;组合E×F、D×F在两次调查中的寒害指数均大于90 %,而组合A×F、B×F、C×F的寒害指数介于65.3 %~94.8 %之间。

表2 甘蓝杂交组合的寒害指数 %
2.3 方差分析
2.3.1 随机区组方差分析 对15个杂交组合的抗寒性进行随机区组方差分析,两个时期的F值分别为130.29和78.92,差异均达极显著水平,表明组合间存在真实的差异,可以进行配合力和遗传力分析。
2.3.2 配合力方差分析 将组合间的方差分解为一般配合力(GCA)方差和特殊配合力(SCA)方差进行分析,两次调查中这两种配合力的F值都达到极显著水平(表3)。这表明,甘蓝抗寒性在两次调查中有极显著的一般配合力差异及组合间极显著的特殊配合力差异;同时还表明,在本试验中基因加性效应和非加性效应对甘蓝抗寒性均起重要作用。
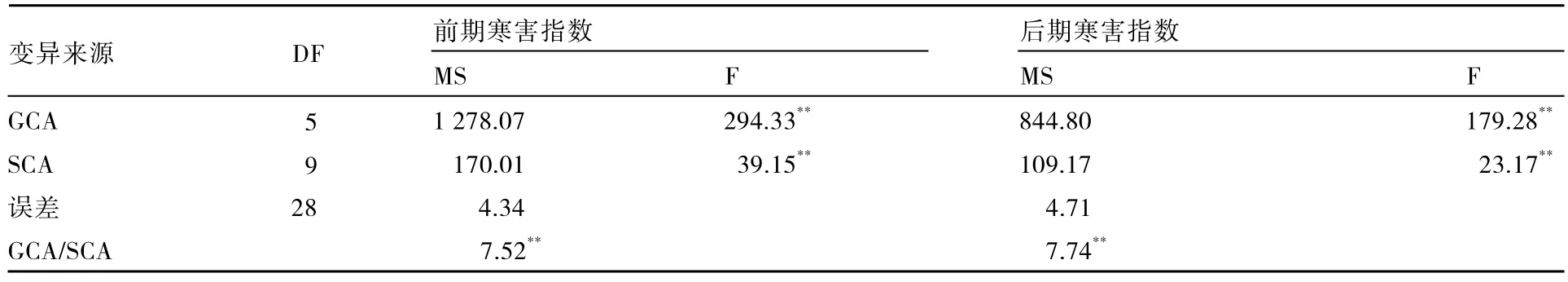
表3 甘蓝杂交组合寒害指数的配合力方差分析
GCA与 SCA均方比是衡量一般配合力和特殊配合力相对重要性的指标。这一比值在两个调查时期均大于 7,且达到极显著水平。由此说明,甘蓝抗寒性主要是由基因加性效应决定的。后期寒害指数的 GCA/SCA均方比略大于前期,这也表明随着温度的降低,基因加性效应的作用略有增加。
2.4 配合力效应及评价
2.4.1 一般配合力效应及评价 一般配合力由基因的加性效应决定,一般配合力高的品系,其相应性状传递能力强,对杂种后代影响也大。由表4可知,在调查的两个时期,自交系F的GCA均为正值,对寒害指数产生正向促进效应。自交系 B、C、D的 GCA均为负值,对寒害指数产生负向促进效应;平均负向效应值绝对值大小顺序为B>D>C,自交系B的GCA效应值最小,杂交组合的寒害指数促进效应最小,即对抗寒性作用最大。因此,自交系 B、C、D的一般配合力效应值为负向,在甘蓝抗寒育种中有很大潜力,可用于试配抗寒组合。自交系A在调查初期表现为正向促进效应,到后期表现为负向促进效应,表明该自交系前期抗寒性略差,待适应零下低温后可诱导出较强的抗寒性。自交系E在调查初期表现为负向促进效应,后期为正向促进效应,表明该自交系的前期抗寒性较好,而后期抗寒性一般。
2.4.2 特殊配合力效应及评价 特殊配合力是指两亲本配制一代杂种的水平,由基因的非加性效应决定。由表5可以看出,不同组合间的SCA效应差异较大。有些组合在前、后期表现出一致的正向或负向优势,如组合A×B、A×C、A×E、B×C、B×D、B×E、B×F、C×D、C×F、D×E、D×F、E×F,在两个调查时期表现出较好的一致性;其他 3个组合的 SCA效应值在两个调查时期正向或负向趋势不完全一致。
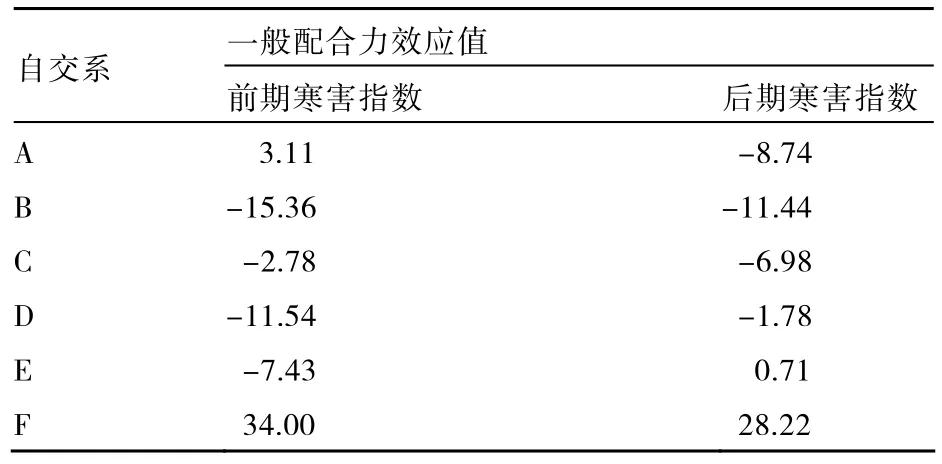
表4 6份甘蓝自交系寒害指数的一般配合力效应值

表5 甘蓝杂交组合寒害指数的特殊配合力效应值
其中组合 A×B、B×F、C×D、C×F、D×E的 SCA效应值在两个调查时期均为负值,抗寒性较强,且表现出很好的一致性。组合D×E的负向SCA效应值最大,其次为组合B×F、A×B,均表现出较好的抗寒性。特别是组合A×B,球色绿,球形好,产量高,后期抗寒性好。
2.5 抗寒性的基因型和表现型方差及遗传力估算
由表6可以看出,在调查的两个时期加性方差均大于非加性方差,表明甘蓝的抗寒性以加性效应为主,还有非加性效应的作用。甘蓝杂交组合在调查的两个时期均表现出较高的遗传力,广义遗传力达97 %以上,狭义遗传力在广义遗传力中所占比例较高,这也表明甘蓝的抗寒性主要以加性效应为主,这与GCA/SCA均方比大于7的结果相符合。

表6 甘蓝杂交组合寒害指数的基因型和表现型方差及遗传力
2.6 高配合力优良抗寒甘蓝组合的选育
综合一般配合力、特殊配合力测定结果及田间经济性状调查结果,初步明确优良抗寒越冬甘蓝组合2个(表7):10秋1119(D×E)、10秋1126(A×B)。10秋1119表现叶色深绿,中心柱长占球高的55 %,结球紧实,结构好,田间抗病性强;10秋1126表现叶色绿,中心柱长小于球高的1/2,结球紧实,结构好,田间抗病性强。

表7 2个优良抗寒甘蓝组合的经济性状
3 结论与讨论
对小麦(孙贵臣和徐环澄,1993)、水稻(Cruz et al.,2010)的研究表明,抗寒性受加性效应和非加性效应共同作用,以加性效应为主。但也有例外,Cruz等(2006)对水稻发芽期的抗寒性配合力研究表明,非加性效应起主要作用,可在较晚的世代进行该性状筛选,推测是因为该抗寒性是在种子发芽期调查的,而Cruz等(2010)调查的抗寒性是在水稻苗期。本试验以6份甘蓝抗寒自交系进行配合力、遗传力分析,结果表明基因加性效应和非加性效应对抗寒性均起重要作用,且以加性效应为主。因此,选择耐寒材料时,应重视从早期世代起对抗寒性状加以筛选。
前人对甘蓝重要性状的配合力测定主要包括耐裂球性(Chiang,1972;庄木 等,2009)、单球质量、中心柱长(方智远和孙培田,1982;More & Wallace,1988)、耐抽薹性(李梅 等,2009)等性状,其中耐裂球性、耐抽薹性、中心柱长、株高等性状的表现以加性效应为主,而单球质量等性状的表现以非加性效应为主。在本试验中,对抗寒性的配合力分析结果表明其以加性效应为主。综合前人研究结果,上述农艺性状中大多数以加性效应为主,表明常规育种经过多代分离筛选,能逐步累积控制优良性状的基因。
在抗寒材料的自然低温筛选过程中,大多数材料的抗寒性表现前后是一致的,且随着温度的降低其加性效应只轻微增加。考虑到多数北方地区冬季极端温度可达-10 ℃以下,因此在实际育种过程中,可以在-3 ℃左右进行抗寒材料的初选,在-5 ℃时做进一步选择,这样既能保证选育材料的成活率,避免受过度冻伤,又达到了抗寒筛选的目的。
方智远,孙培田.1982.甘蓝自交系几个数量性状配合力的分析初报.中国农业科学,15(1):49-55.
方智远.2008.我国甘蓝产销变化与育种对策.中国蔬菜,(1):1-2.
李梅,刘玉梅,方智远,杨丽梅,庄木,张扬勇,孙培田,孙振英.2009.结球甘蓝自交系抽薹与开花性状配合力及遗传力分析.华北农学报,24(5):86-89.
李锡香,方智远.2007.结球甘蓝种质资源描述规范和数据标准.北京:中国农业出版社.
刘来福,毛盛贤,黄远樟.1984.作物数量遗传.北京:农业出版社.
孙贵臣,徐环澄.1993.冬小麦品种抗寒性遗传研究.山西农业科学,21(4):4-7.
杨丽梅,方智远,刘玉梅,庄木,张扬勇,孙培田.2011.“十一五”我国甘蓝遗传育种研究进展.中国蔬菜,(2):1-10.
庄木,张扬勇,方智远,刘玉梅,杨丽梅,孙培田.2009.结球甘蓝耐裂球性状的配合力及遗传力研究.中国蔬菜,(2):12-15.
Chiang M S.1972.Inheritance of head splitting in cabbage(Brassica oleracea L.var.capitata L.).Euphytica,21(3):507-509.
Cruz R P,Duarte I T L,Cabreira C.2010.Inheritance of rice cold tolerance at the seedling stage.Scientia Agricola,67(6):669-674.
Cruz R P,Milach S C K,Federizzi L C.2006.Inheritance of rice cold tolerance at the germination stage.Genetics and Molecular Biology,29(2):314-320.
More T A,Wallace D H.1988.Combining ability and heterisis studies using selfincompatible lines of cabbage.Indian Journal of Genetics,47:20-27.
猜你喜欢
蔬菜(2022年10期)2022-11-10
养猪(2022年4期)2022-08-17
今日农业(2021年20期)2021-11-26
河北果树(2020年4期)2020-11-26
河北果树(2020年1期)2020-02-09
中国种业(2018年11期)2018-11-16
猪业科学(2018年5期)2018-07-17
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
园艺与种苗(2015年8期)2015-12-28
