
开化王山采种基地森林群落数量分类与多样性研究
2011-05-08 07:08:02汪良义程清明陈声文唐群儿陈建华
浙江林业科技 2011年6期
贾 琪,方 腾,汪良义,程清明,陈声文,唐群儿,陈建华*
(1. 浙江师范大学化学与生命科学学院,浙江 金华 321004;2. 浙江开化古田山国家级自然保护区,浙江 开化 324306;3. 开化县林业开发公司,浙江 开化 324300)
开化王山采种基地森林群落数量分类与多样性研究
贾 琪1,方 腾2,汪良义3,程清明3,陈声文2,唐群儿1,陈建华1*
(1. 浙江师范大学化学与生命科学学院,浙江 金华 321004;2. 浙江开化古田山国家级自然保护区,浙江 开化 324306;3. 开化县林业开发公司,浙江 开化 324300)
在浙江开化王山阔叶林采种基地群落学调查基础上,根据每个样地内重要值大于1.0的102种木本植物的具体数据,对31个样地群落类型进行聚类分析,并对各样地木本植物的物种多样性和物种丰富度指数进行分析。结果表明:(1)王山采种基地木本植物群落类型主要可分为甜槠林、拟赤杨林、短尾柯林、乌楣栲林、木荷林、马银花林、红楠林、钩栲林、杉木林9个类型,总体以常绿落叶林为主,也有少量常绿落叶阔叶混交林和人工杉木林。(2)物种多样性指数(Shannon-Weiner 指数)和物种丰富度整体表现出相似趋势,即乔木层>灌木层;不同群落类型中乔灌两层的多样性指数有一定差异。
王山;群落类型;聚类分析;物种多样性;物种丰富度
随着天然阔叶林逐渐减少,建立采种基地以保护和繁育优良种质树种已成为林业建设的重点之一[1~2]。浙江开化王山阔叶林采种基地是浙江省林业种苗管理总站和开化县林业开发公司利用原有的天然常绿阔叶林建成的,对研究和保护珍稀树种、提高常绿阔叶林种质资源产量具有非常积极的意义,程清明等对王山常绿阔叶林群落的结构特点已有一定的研究[3],但对王山采种基地森林群落具体类型及物种多样性特征等方面缺乏具体研究。本文通过等级聚类分析方法,量化分析王山采种基地森林群落的基本类型、样地内木本植物的物种多样性等特点,较为全面地分析王山阔叶林采种基地植被基本现状,对王山基地群落生物多样性的成因和演替趋势作出初步分析,以期对今后的相关研究提供一定的理论依据。
1 自然概况与研究方法
1.1 自然概况
开化王山阔叶林采种基地位于浙江省开化县杨林镇川南村王山,面积1 470 hm2,现有森林蓄积量5.2万m3,其中阔叶林面积占75%。基地属南岭山系怀玉山脉,主峰千思坦,海拔997.4 m,分王山和南华山小圬两块,各有1条大沟和许多支流,低山沟谷两侧悬崖裸露地形陡峭,中高山地形逐渐趋向缓和,整个山峰呈西南-东北走向,水流注入衢江汇于钱塘江。母岩以花岗岩为主,土壤肥厚,主要类型有红壤、红黄壤、黄红壤。
王山阔叶林采种基地属我国亚热带季风气候区,受季风影响大,四季分明,雨量充沛。年平均温度15.3℃,生长期总积温5 125.4℃,无霜期约250 d,年均降水量1 762.1 mm,相对湿度为81%。从2003年到2004年林场对位于基地中心的红滕湾一带进行了一定程度的抚育,抚育面积为767 hm2,包括共12个林班56个小班,海拔在400 ~ 800 m。抚育方式以清除林下灌草和枯立木、病死木为主[3]。
1.2 研究方法
1.2.1 样方设置 群落学调查始于2005年10月。首先对基地中12个林班进行全面踏查,记录每个林班的起源、地貌、群落特征、海拔、土壤、周围环境和外围影响,对群落结构进行一般描述;然后在每个林班选择具有代表性的样地1 ~ 4个。共设置了31个样地。样地面积均为20 m×20 m。
1.2.1 样地调查 将样地划分为16个5 m×5 m的样方,每间隔一个样方的右下角划出2 m×2 m的小样方调查灌木层(包括幼树、幼苗),用同样方法在2 m×2 m的小样方内划出1 m×1 m的小样方调查草本层。对胸径≥1 cm的乔木类植物逐株调查,记录种名、胸径、高度、枝下高、冠幅等,灌木层和草本层记录种名、株数、高度、数量、盖度等,并记录层间植物。
1.2.2 数据统计分析 聚类分析采用了等级聚类方法,距离矩阵选择了Bray-curtis距离。用Past软件进行聚类分析计算,用R和Excel等软件进行多样性指标的计算。
乔木层重要值为(相对多度+相对频度+相对显著度)/3,灌木层及草本层重要值为(相对频度+相对盖度)/2[3]。
根据物种多样性测度指数应用的广泛程度以及对群落物种多样性状况的反映能力,本文选取以下2种多样性指数来测度和分析群落物种多样性特征,并以重要值Pi作为多样性指数的测度依据。
物种丰富度指数(S)为样地中物种总数。
Shannon-Wiener指数[4~5]式中,Pi为种i的相对重要值,S为样地中出现的植物种类数量。
2 结果与分析
2.1 群落类型分析
生物群落分类是生态学研究领域中争论最多的问题之一,植物群落分类的基本单位是群丛(association)[6]。目前用数学方法对样地植被资料进行分析归类已经受到了普遍的重视,因为用这种方法进行分类可以获得较为客观的结果,在群落类型的分类上,聚类分析法优于优势种划分法[5~6]。本文以开化王山阔叶林采种基地不同样地木本层重要值大于1的102个物种为对象,对31个样地进行聚类分析(图1)。

图1 王山阔叶林采种基地群落类型聚类分析Figure 1 Dendrogram of stand similarities of 31 plots in Wangshan
从聚类的结果看,王山阔叶林采种基地群落类型大致可划分为9组,总体以常绿落叶林为主,也有少量常绿落叶阔叶混交林和人工杉木林,具体如下:
2.1.1 甜槠林 包括1、2、3、10、19、20、27共7个样地。以甜槠(Castanopsis eyrei)为优势种的常绿阔叶林在中国中亚热带东部常绿阔叶林中具有典型性和代表性[7],王山采种基地的甜槠林主要分布在海拔350 ~ 750 m的山坡和山地沟谷。物种组成以壳斗科、樟科、山茶科为主。林冠郁闭度大,乔木层盖度在80%左右。乔木层可以分为两个亚层,第一亚层优势种为甜槠,重要值19 ~ 36,虎皮楠(Daphniphyllum oldhami)、木荷(Schima superba)、丝栗栲(C. fargesii)、红楠(Machilus thunbergii)、杜英(Elaeocarpus sylvestris)等常绿阔叶树种占据很大比例,重要值常常超过10。第二亚层中除了以上优势树种外,有较多的鹿角杜鹃(Rhododendron latoucheae)、马银花(R. ovatum)等小乔木。灌木层以乔木层小苗及矩形叶鼠刺(Itea chinensisvar.oblonga),
山矾(Symplocos caudata)、刚竹(Phyllostachys viridis)、细枝柃(Eurya loquaiana)、毛花连蕊茶(Camellia fraterna)、黄瑞木(Adinandra millettii)、薯豆(Elacocarpus japonicus)的幼苗为常见种。草本层以狗脊蕨(Woodwardia japonica)、苔草(Carexspp.)、里白(Diplopterygium glaucum)最为多见。根据次优势种的不同,可将甜槠林细分为甜槠—木荷林(如样地1和27)、甜槠—丝栗栲林(如样地2和3)、甜槠—虎皮楠林(样地10、19、20)3种类型。
2.1.2 拟赤杨林 拟赤杨林在王山采种基地也较多,包括7、8、15、16、17、18、26共7个样地。从海拔450 ~ 850 m的沟谷地带均有分布。乔木层盖度较小,在60%左右。物种组成复杂,乔木层优势种不十分明显,其中落叶树种总体以拟赤杨(Alniphyllum fortunei)占优势,平均重要值达12.5,檫木(Sassafras tzumu)也较多,平均重要值达7.9,另含一定数量的野漆树(Toxicodendron succedaneum)、山桐子(Idesia polycarpa)、山合欢(Albizzia kalkora)等;常绿植物则以薯豆、红楠(Machilus thunbergii)、丝栗栲、虎皮楠、甜槠等为主,重要值平均在10左右,故本类型主要属常绿落叶阔叶混交林,其中16和17号样方有一定数量的杉木(Cunninghamia lanceolata),有人工种植干扰痕迹。灌木层以乔木层小苗及杜茎山(Maesa japonica)、浙江新木姜子(Neolitsea auratavar.chekiangensis)、鹿角杜鹃、细枝柃、乌药(Lindera aggregata)、檵木(Loropetalum chinensis)为优势。苔草、淡竹叶(Lophatherum gracilie)、里白、阔叶山麦冬(Liriope platyphylla)在草本层中为优势种。
2.1.3 乌楣栲林 分布于400 ~ 800 m的山坡上,包括11、12、13、21共4个样地。乔木层平均盖度70%,优势种主要为乌楣栲(Castanopsis jucunda),平均重要值达16.7,青冈、石栎(Lithocarpus glabra)、虎皮楠、木荷的平均重要值在6.3 ~ 9.1,拟赤杨、细叶青冈(Cyclobalanopsis myrsinaefolia)、野漆树等也有少量分布。灌木层以乔木层小苗及格药柃、毛花连蕊茶(Camellia fraterna)、杜茎山等为主。草本层盖度较小,分布有苔草、狗脊蕨等。层间植物有络石(Trachelospermum jasminoides)等。
2.1.4 短尾柯林 短尾柯林分布于海拔600 ~ 700 m的山谷和山坡上,包括4、22、25、30、31共5个样地。乔木层盖度70%左右,优势种为短尾柯(Lithocarpus harlandii),平均重要值15.3,红楠(平均重要值8.9)、拟赤杨(4.8)、青冈(3.2)、木荷(2.5)、锥栗(Castanea henryi)(2.0)、甜槠(1.8)均有少量分布。灌木层以乔木层小苗及油茶、朱砂根(Ardisia crenata)、太平莓(Rubus pacificus)等为主。草本层有金星蕨(Parathelypteris glanduligera)、麦冬(Ophiopogon japonicus)等,层间植物以香花崖豆藤(Millettia dielsiana)等为主。
2.1.5 木荷林 有23、24号两个样地,位于海拔650 ~ 950 m的山坡上,林冠盖度70%。样地种以木荷为绝对优势种,平均重要值达31.2,甜槠次之,平均重要值为11.5,其他如红楠、豹皮樟(Litsea coreanavar.sinensis)、锥栗、冬青等也有一定分布,平均重要值在2.0 ~ 5.0,值得注意的是本样地有较多的小乔木鹿角杜鹃,平均重要值达13.9。灌木层以乔木层小苗及阔叶箬竹(Indocalamus latifolius)、油茶、太平莓为主。草本层常见苔草、金星蕨、阔叶山麦冬等。
2.1.6 马银花林 分布于海拔700 ~ 800 m的山坡上,有28和29两个样地。林冠层盖度很差或缺少高大乔木。亚乔木层郁闭度很大可高达80%,小乔木马银花为最优势物种,平均重要值为21.8,其次为格药柃(13.6),甜槠、木荷、短尾柯、细叶青冈、锥栗等乔木仍有一定数量分布,平均重要值在4.4 ~ 8.6。灌木层以乔木层小苗及小叶青冈(Cyclobalanopsis myrsinifolia)等为主。层间植物和草本层盖度很小,有少量的络石、狗脊等。
2.1.7 杉木林 分布于海拔350 ~ 600的山坡上,林冠郁闭整齐,盖度高达70%,包括5、6两个样地。杉木为人工栽培,占据绝对优势,平均重要值高达55.3。拟赤杨等天然起源,平均重要值达19.2,也占有重要地位,此外檫木、盐肤木(Rhus chinensis)、山鸡椒(Laurus cubeba)、垂珠花(Styrax dasyanthus)、小果冬青(Ilex micrococoa)等落叶树种也有一定数量的分布,平均重要值在1.0 ~ 3.3。灌木层以乔木小苗及格药柃(Eurya muricata)为优势种,草本层有苔草、狗脊蕨等。
2.1.8 红楠林 仅9号样地。位于海拔470 m左右的沟谷及两侧,乔木层盖度70%,为50 a左右的常绿阔叶林。优势种为红楠,重要值18.4,虎皮楠、南酸枣、山合欢等高大乔木也有一定数量分布,平均重要值在4.2 ~ 8.9,亚乔木层油茶最为优势,重要值达10.8,其次是马银花(6.0)、山矾(4.7)等。灌木层有乔木小苗及狗骨柴(Diplospora dubia)、细枝柃、矩形叶鼠刺等。草本层盖度较低,以芒萁(Dicranopteris dichotoma)为主。
2.1.9 钩栲林 仅14号样地。位于海拔550 m的山地沟谷两侧。乔木层盖度非常高,达90%,优势种钩栲(Castanopsis tibetana)的重要值是29.5,次优势种拟赤杨重要值13.0,分布有少量的虎皮楠(6.2)、小果冬青(5.8)、青钱柳(3.8)、蓝果树(3.4)等。灌木层以乔木层小苗及杜茎山等最多。草本盖度较高,有华东瘤足蕨(Plagiogyria japonica)、狗脊、淡竹叶等,层间植物有络石等。2.2 群落物种多样性
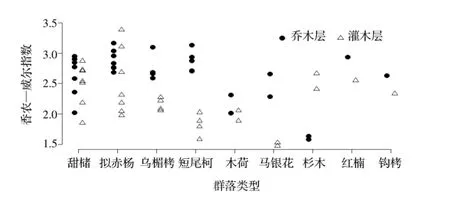
图2 样地乔木层与灌木层物种多样性指数Figure 2 Shannon-wiener index of canopy and shrub layer
森林生物多样性在一定程度上是衡量森林质量的重要指标,丰富的物种多样性是森林生态系统稳定的重要标志[8~9]。分别考察聚类分析产生的9类群落类型的生物多样性(Shannon-Weiner 指数),整体表现为乔木层 >灌木层(图2),这与之前的研究结果基本一致[3]。通常情况下,森林群落中灌木层的物种多样性较高。但是王山采种基地灌木层的多样性整体上明显小于乔木层,可能与2003-2004年林下灌草的清除有很大关系。从具体群落类型看,常绿阔叶林的乔木层生物多样性指数普遍大于灌木,不同的群落类型中乔灌两层的生物多样性指数有一定差异,甜槠林是典型的亚热带常绿阔叶林型,在王山分布面积最广,乔灌层的多样性都维持在较高的水平;其他常绿阔叶林如乌楣栲林、短尾柯林、钩栲林、红楠林等都具有较高的乔木物种多样性和较低的灌木物种多样性;木荷林分布于高海拔山坡上,乔木和灌木多样性都较低;马银花林由于林冠低矮、盖度大,林下灌木、草本生长受到限制,灌木层多样性特别低。常绿落叶阔叶混交林和杉木林的情况相对复杂,王山的拟赤杨林属于次生演替的中后期,包含较多的阳生性落叶或常绿阔叶树种,乔木层物种多样性很高,灌木层受林下干扰,多样性变异较大。杉木人工林灌木丛层多样性较高,但乔木层多样性指数很低,这可能和自然演替时间较短有关,上层物种主要为杉木,下层物种还多未到达乔木层[10]。
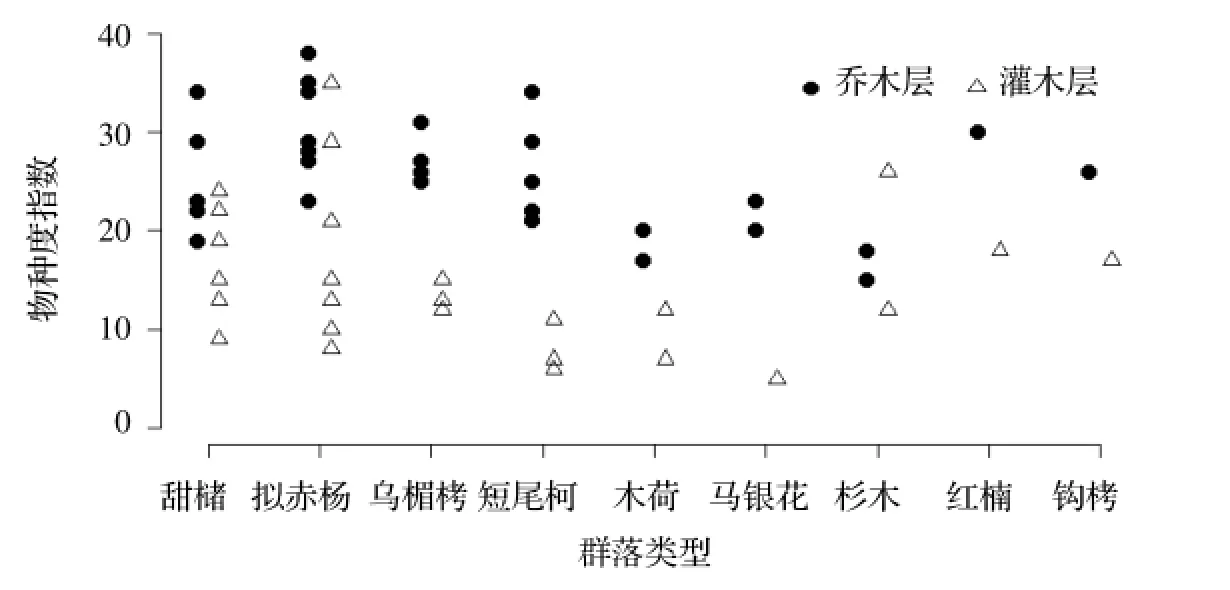
图3 样地乔木层与灌木层物种丰富度指数Fig ure 3 Sp ecies richness of canopy and shrub layer
物种丰富度指数(如图3)和shannon-weiner指数具有相似的规律,只有杉木林稍有不同,杉木林乔木层的shannon-weiner指数非常低,仅为1.6,但物种丰富度指数却几乎达到了木荷林的水平。
3 结论与讨论
(1)以开化王山阔叶林采种基地不同群落木本植物重要值大于1的120个物种为考察对象,对31个样地进行聚类分析显示,王山森林群落基本上可以分为9个类型,具体包含甜槠林、拟赤杨林、短尾柯林、乌楣栲林、木荷林、马银花林、红楠林、钩栲林、杉木林等,总体以常绿落叶林为主,也有少量常绿落叶阔叶混交林和人工杉木林。甜槠林是一类典型的亚热带常绿阔叶林,属于演替顶级群落[11],拟赤杨材可能是优势的阔叶落叶混交林遭遇砍伐干扰后,在保护较好的情况下演替发展起来的[12],而杉木林为人工林,处于演替的早期。可见采种基地内群落类型比较丰富,涵盖了几乎所有的演替阶段,这为适合不同演替阶段的物种的共存创造了良好的条件,十分有利于高物种多样性的维持和稀有珍稀物种的保护。
(2)王山森林木本植物多样性指数(Shannon-Weiner 指数)和物种丰富度整体表现为乔木层 > 灌木层。这可能与采种基地进行有目的林下灌草清理等抚育活动有关系。不同的群落类型,乔灌两层的shannon-weiner指数和物种丰富度指数都有一定差异,如甜槠林、乌楣栲林、短尾柯林、红楠林、钩栲林以及大部分的拟赤杨林都相对具有较高的乔灌层shannon-weiner指数和物种丰富度指数;木荷林分布于高海拔山坡上,乔木和灌木两种多样性都较低。人工杉木林虽然有很低的乔木层shannon-weiner指数(平均1.5),但是物种丰富度仍达16.5,和木荷林(18.5)基本持平,原因是人工杉木林乔木层中除杉木植株数量较多外,其他物种种类比预期的要多,但植株数量较少,另一方面也反映出尽管人工杉木林中杉木占绝对优势,但本地植物仍可通过各种途径扩增或繁衍。
(3)王山采种基地在历史条件和人为抚育管理的情况下,具有丰富的群落组成类型和结构类型,生物多样性较高。不同的森林群落类型,其物种组成上存在较大差异。从采种基地的角度来看,这有利于保存更多的物种,但是自然状态下的演替会使得一些物种增加,而另一些演替早期的物种尤其是落叶树种会处于弱势甚至消失[13]。对不同的林型需要编制不同的营林方案,确定小班内的主要目的树种,清除非目标树种,如以拟赤杨等落叶树种为主的常绿落叶阔叶混交林或人工杉木林等,需要定期择伐抚育以保证其良好更新,而一些常绿阔叶树种仅需要让其自然更新,至于抚育的具体力度、频率和抚育方式的选择上需结合森林的实际情况和采种基地的目标进行进一步的探讨。
致谢:感谢浙江省林业种苗管理总站对开化王山采种基地的大力支持!
参考文献:
[1] 蔡勇. 福建省延平区林木采种基地项目的建设方案[J]. 安徽农学通报,2007,13(10):114-115.
[2] 李军. 临海市阔叶树采种基地营建与管理技术探析[J]. 浙江林业科技,2006,26(4):46-49.
[3] 程清明,汪良义,方腾,等. 开化王山阔叶林采种基地森林群落结构研究[J]. 浙江林业科技,2006,26(4):19-23.
[4] 马克平. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J]. 生物多样性,1994,2(3):162-168.
[5] 戈峰. 现代生态学[M]. 北京:科学出版社,2008.
[6] 孙儒泳. 基础生态学[M]. 北京:高等教育出版社,2002.
[7] 胡正华,于明坚,索福喜. 古田山国家自然保护区常绿阔叶林植物物种多样性研究[J]. 中国农学通报,2005,21(3):134-137.
[8] 张庆,牛建明,王秀梅. 生物多样性与生态系统功能关系研究进展[J]. 生物学通报,2009,44(1):15-17.
[9] 翀张步,李凤民,黄高宝. 生物多样性对生态系统功能及其稳定性的影响[J]. 中国生态农业学报,2006,14(4):12-15.
[10] 林开敏,张文富,谢国阳,等. 老龄杉木林下天然更新阔叶植被物种多样性研究[J]. 福建林学院学报,1997,17(4):313-317.
[11] 李立,陈建华,任海保,等. 古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J]. 植物生态学报,2010,34(3):241-252.
[12] 王献溥,李俊清. 广西拟赤杨林分类和演替的研究[J]. 植物研究,2000,20(3):332-350.
[13] 李翠环,余树全,周国模. 亚热带常绿阔叶林植被恢复研究进展[J]. 浙江林学院学报,2002,19(3):101-105.
Community Type, Size Distribution and Species Diversity Analysis of Kaihua Wangshan Seed Base
JIA Qi1,FANG Teng2,WANG Liang-yi3,CHENG Qing-ming3,CHEN Sheng-wen2,TANG Qun-er1,CHEN Jian-hua1*
(1. College of Chemistry and Life Science, Zhejiang Normal University, Jinhua 321004, China; 2. Gutianshan National Nature Reserve, Kaihua 324306, China; 3. Kaihua Forestry Exploitation Company, Kaihua 324300, China)
Cluster method is used to classify 31 plots via analyzing the importance value of 102 tree species, while biodiversity analysis are also used to clarify the development status of these forest plots. The results showed that: (1) Communities of Wangshan forest can be classified into nine groups, the most dominant species are Castanopsis eyrei, Alniphyllum fortunei, Lithocarpus harlandii, Castanopsis jucunda, Schima superba, Rhododendron ovatum, Machilus thunbergii, Castanopsis tibetana and Cunninghamia lanceolata respectively, most of them are belonged to evergreen broad-leafed forest, some of them are mixed evergreen & deciduous forest or man-made forest. (2) Species diversity and species richness indexes are larger in tree layer than that in ground layer and varied between different forest community types. This study will give advanced understanding in the status of Wangshan forest and provide some reference on forest management.
Wang Shan, Community type, Cluster analysis, Species diversity, species richness
S718.54
A
1001-3776(2011)06-0018-06
2011-04-26;
2011-08-16
浙江省自然科学基金“密度制约效应对古田山常绿阔叶林生物多样性影响的研究”(Y5100361)资助
贾琪(1985-),男,陕西靖边人,硕士生,从事种子植物多样性研究;*通讯作者。
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
绿色中国(2019年18期)2020-01-04 01:57:08
乡村科技(2018年8期)2018-02-13 12:16:18
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:55
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
湖北农业科学(2013年6期)2013-04-29 14:29:32
