
重庆市桃花溪受损河岸植被恢复初期物种多样性变化及其对土壤环境效应的影响
2011-05-07 11:06:56孟凡超王玉杰赵占军赵洋毅雷声坤
水土保持研究 2011年4期
孟凡超,王玉杰,赵占军,赵洋毅,雷声坤
(北京林业大学水土保持学院水土保持与荒漠化防治教育部重点实验室,北京100083)
物种多样性是一个群落结构和功能复杂性的度量,表征着生物群落和生态系统的结构复杂性,是揭示植被组织水平的生态学基础[1]。国内外大多涉及到的领域包括森林群落[1-3],草原群落和人工草地群落[4-5],退化沙质草地和盐碱沙地群落[6-8],湿地植物群落[9]以及不同树种搭配及空间配置下水源涵养林[10]的物种多样性及其土壤特征关系的研究,对于河岸过渡带的物种多样性及其影响因素则研究较少。如今,物种多样性已经成为许多受损河岸健康评价的重要指标之一[11],揭示不同护岸模式物种多样性的特征,阐明物种多样性与土壤因子的相关性,为受损河岸物种多样性的保护与持续利用提供理论依据已经尤为必要。
河岸生态护坡作为一种“无阻隔、无污染、无副作用”的受损河岸治理技术已经被广泛应用,它对于重建受损河岸生态系统,恢复固坡、截污等生态功能,保持水陆生态系统的连贯性具有重要意义[12]。在重庆市长寿区桃花溪,部分河岸段由于受到较为频繁的人为干扰,特别是开发建设项目弃渣堆积,护岸植被严重破坏,植物多样性较其他河岸段锐减,地表裸露严重。因此,选取适宜的河岸修复技术,利用河岸土壤作为植物生长的重要物质基础,研究其物理性质、化学性质和土壤质地的差异对河岸植被的物种多样性的影响[13],并通过对生物多样性进行定期或不定期的监测,确定与预期标准相一致或相背离的程度,监测治理初期是否达到受损河岸生态修复的目的[14]。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于重庆市长寿区桃花溪,长寿区位于重庆东部 ,地处 106°49′-107°27′E ,29°43′-30°12′N ,属中亚热带湿润气候区,四季分明、气候温和、冬暖春早、热量丰富、降水充沛、伏旱多雾,常年平均气温17.7℃,最高年20.4℃,最低年16.7℃,常年平均降水量1 165.7 mm。辖区南北长 56.5 km,东西距57.5 km,总面积1 423.62 km2。属重庆市三峡库区生态经济区29个区县之一,处于三峡库区生态经济区和都市发达经济圈的结合部。
长寿境内河流众多,水资源较为丰富,工农业生产比较发达,素有“化工城”之称。流域面积361.8 km2,全长 63.1 km ,天然落差 914.2 m,平均比降8.08‰,多年平均流量5.3 m3/s。受损河岸示范区域位于桃花溪左岸一级阶地平台上,在长久的桃花溪洪水的不断冲刷下,河漫滩已变成弧形,河岸土壤属侏罗系沙溪庙组沙泥岩发育而成的灰棕紫泥,呈微碱性。在受损河岸示范区各观测带结合土壤生物工程的理念,采用了抛石结合植物扦插、自然原型护岸、土工织物扁袋结合扦插(织物扁袋可选用麻袋布)、天然材料织物垫4类护坡模式,引进适应性强的乡土植物,结合河岸消落带固有的原生态植被,形成以毛竹-灌草混交的株间混交植物群落。

表1 不同河岸治理模式下岸坡基本概况
1.2 研究内容及方法
1.2.1 野外调查方法 在受损岸坡上,选取面积为15 m×40 m的区域进行常规群落学调查,共设置5个样地,对每个样地的灌木、草本植物进行逐一调查。其中,乔木分成5 m×5 m的小样方若干个,对所有乔木进行每木检尺,调查种名、胸径、树高、冠幅等,并记录乔木数量及郁闭度;灌木层及草本层的调查样方则分别设置为2 m×2 m、1 m×1 m,记录每个种的名称、多度、高度和基径。另外,每个样地的海拔、坡向、坡度和坡位也需详细记录。
桃花溪原生态受损河岸植被稀疏,仅零星分布少量的草本植物,覆盖度仅为30%,包括棒头草(Polypogon f ugax Nees ex Steud)、双穗雀稗(Paspalum paspaloides(Michx.)Scribn)、扁穗牛鞭草(Hemarthria compressa(Linn.f.)R.Br)等。经过生态修复,外来引进适合当地的植物种进行栽植,乔木包括毛竹 、香樟(Cinnamomum camphora(L.)Pres)、垂柳等;灌木包括丁香、黄花槐、冬青、红叶李、木芙蓉等;草本植物包括鸢尾、黑麦草(Lolium perenne Linn)、苏丹草(Sorghum sudanense(Piper)Stapf)等。经过修复治理,桃花溪受损河岸的物种数由原来的3种增加到25种,其中乔木2种,灌木8种,草本15种,生物多样性大大增加。
1.2.2 土壤理化性质测定 样地内,在0-30 cm土壤剖面上按“S”形取样法随机挖取混合土样,每个样地重复3次,采用环刀法测定土壤容重、毛管孔隙度和非毛管孔隙度,开氏消煮法测定全氮,碱解扩散法测定有效氮,碱熔融-钼锑抗比色法测定全磷,钼锑抗比色法(0.03 mol/L NH4F-0.025 mol/L HCl浸提)测定速效磷,碱熔融-火焰光度计法测定全钾,火焰光度法(1 mol/L NH4OAC浸提)测定速效钾,pH 213台式酸度离子计测定pH值,重铬酸钾法测定土壤有机质,乙酸钠-火焰光度法测定阳离子交换量。
1.2.3 多样性指数的测定
(1)重要值的计算方法
乔木重要值=(相对密度+相对优势度+相对频度)/3灌草重要值=(相对多度+相对盖度+相对频度)/3
(2)多样性指数计算。本文选用目前使用较多的4个物种多样性计算模型:Margalef丰富度指数(R)、Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)、Pielou均匀度指数(E)来计算不同生态河岸修复模式的物种多样性[15]。各指数公式如下[15-16]:

式中:S——各群落的物种数目;Ni——群落中某一层次第i个物种的重要值;N——该层次所有物种重要值之和。将群落各层次多样性指数加和得到群落物种总体多样性指数。
1.2.4 数据处理方法 运用SPSS 16.0、Excel统计分析软件对不同河岸修复模式的土壤理化因子进行One Way ANOVA方差分析,同时用一般相关分析与逐步回归(Stepwise)方法分析不同修复模式下物种的多样性指数与土壤理化性质之间的关系。
2 结果与分析
2.1 不同护岸模式下物种多样性指数分析
桃花溪受损河岸示范段是以各种护岸材料结合植被进行的综合护岸模式,选取的植被以灌木或小乔木结合表土固结能力强的草本植物为主,高大乔木栽植较少,因此乔木的多样性较小。在桃花溪受损河岸示范区,经过3 a的植被恢复,河岸带植被群落结构逐渐成型,植被功能趋于完善,主要的优势种垂柳、红叶李、木芙蓉、黄花槐、黑麦草等在垂直层面构建的植被体系最大程度的提高了岸坡的物种多样性水平和水土保持的能力。因此,综合量度各多样性指标显得尤为必要。各岸坡生物多样性指数特征值见表2。

表2 各岸坡不同植被层次多样性指标特征值
乔木层中,由于修复前河岸带没有乔木,因此原生态半裸地乔木层的各多样性指数均为0。在示范区各岸坡修复治理后,只有植被护岸以毛竹和垂柳等乔木种为优势种,其他护岸模式由于以灌草护岸模式为主,其乔木层的多样性几乎为0。其中,仅有天然材料织物垫和人工扁袋护岸栽植了少量的单一垂柳,其Margalef丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数均接近于0,Simpson优势度指数仅为0.972。在植被护岸中,R,H,D,E指数大小分别为 3.476,0.679,1.105,0.98,显示了乔木数量及密度相对较大,且分布较为均匀,但是物种数相对较低,多样性较差。
灌木层中,原生态半裸地的各多样性指数为0。在新增护岸模型中,天然材料织物垫的Margalef丰富度指数最高,达到了14.24,是其它样地的3~4倍,而植被、人工扁袋和抛石护岸的丰富度指数则相差不多。Shannon-Wiener多样性指数大小植被(1.605)>天然织物垫(1.532)>人工扁袋(1.383)>抛石护岸(1.09),差异性不是十分明显,尤其是植被护岸和天然材料织物垫护岸的多样性相比其他岸坡更丰富。Simpson优势度指数大小为植被(1.706)>人工扁袋(1.48)>抛石(1.29)>天然织物垫(1.226),植被护岸明显大于其它岸坡,生态优势度是天然织物垫的1.4倍。抛石、植被、天然材料织物垫的Pielou均匀度指数基本相同,均在0.99以上,天然材料织物垫也达到了0.952,说明各护岸模式灌木林的分布非常均匀。
草本层中,5种护岸模式的Margalef丰富度指数(R)、Shannon-Wiener多样性指数(H)变化规律基本一致,各新增护岸模式的多样性远比半裸地要好,其中,植被护岸的多样性最好,分别达到了11.679和2.29,是半裸地的3倍左右,其草本物种数和个体数量最大。Simpson优势度指数中天然材料织物垫(2.767)>植被(1.895)>抛石(1.704)>半裸地(1.294)>人工扁袋(1.079),天然织物垫、植被、抛石护岸的优势度明显高于半裸地与人工扁袋。而Pielou均匀度指数基本相同,高达0.97以上。
在总体群落中,植被护岸的植物生长最好,物种丰富,分布较均匀,物种多样性指数较高。此护岸模式下提高物种多样性作用极显著,效果最好。从多样性指数变化规律上看,各护岸模式的多样性指数特征值与草本层的特征值相关性最高,变化规律趋于一致,说明在总体群落中,草本层的物种数和数量占据绝对优势,成为影响总体群落的最主要层次。
2.2 不同护岸模式的土壤特征因子分析
土壤是植物生长的基质,其理化特性决定着植物群落类型的分布,同时植物群落在整个生长发育过程中也能通过根系分泌物和枯落物等改善土壤的水、热、气、肥等理化性质[17]。土壤含水率、土壤容重、毛管孔隙度和非毛管孔隙度等土壤物理指标是土壤持水性能的重要体现,综合反映了土壤的透水、持水能力和基本物理性能[18]。从土壤保水性能来看,毛管孔隙中的水可长时间保存在土壤中,主要用于植物根系吸收和土壤蒸发,而非毛管孔隙中的水可以及时排空,更有利于水分的下渗,同时保持土壤更高的通气性[19]。pH 值、有机质含量 、全氮、全磷、全钾、速效氮、有效磷、速效钾和阳离子交换量(CEC)等土壤化学指标综合表征了土壤养分的状况,对植物生长提供必需的营养物质,同时促进土壤的团粒结构,增加黏粒与土壤保水保肥的特性,更有利于植物根系在土壤中的生长延伸。因此,土壤理化性质对生物多样性有一定程度的影响,对各理化指标的分析显得尤为必要。
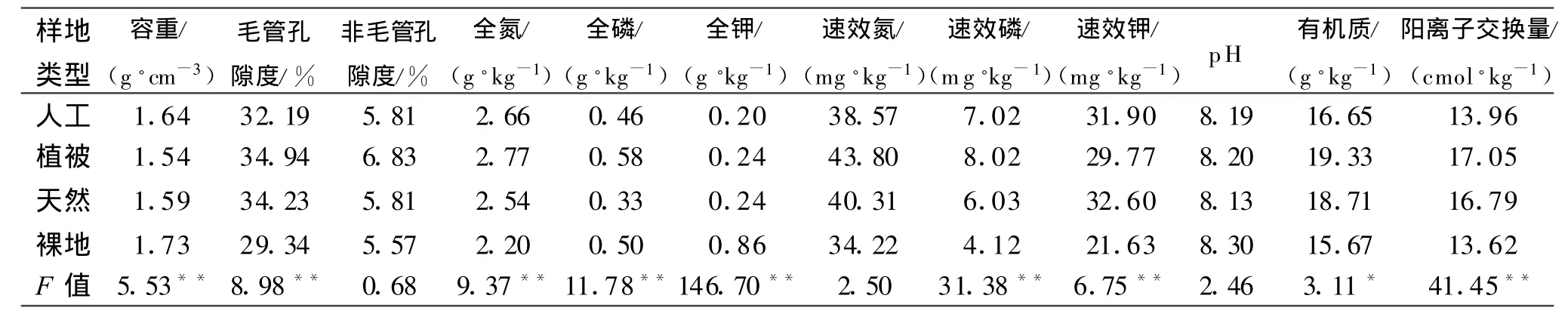
表3 各岸坡土壤特征因子平均值及差异值表
如表3所示,对不同护岸模式的土壤特征因子进行均值及方差分析,发现各岸坡较对照区的容重、毛管孔隙度 、全N 、全P、全K、速效P、速效K 和CEC呈极显著性差异,有机质呈显著性差异,而非毛管孔隙度、速效N、pH差异性不显著,说明各受损岸坡的治理模式对土壤的大多数理化性指标有极显著的影响。从物理性指标上分析,植被护岸和天然材料织物垫护岸的容重小于原生态的半裸地,但是其土壤总孔隙度与毛管孔隙度最大,其次是人工织物扁袋,最差的是半裸地。其中,毛管孔隙大小为植被(34.94%)>天然织物垫(34.23%)>人工扁袋(32.19%)>半裸地(29.34%),且差异性极显著(F=8.98;P<0.01);非毛管孔隙度大小为植被(6.83%)>天然织物垫(5.81%)>人工扁袋(5.81%)>半裸地(5.57%),但差异性极小(F=0.68)。说明植被护岸与天然材料织物垫对土壤的保水性与持水性最优,而半裸地最差,但各护岸措施对土壤大孔隙的通气性能改善效果不明显。从化学性指标上分析,植被护岸与天然材料织物垫护岸在有机质、CEC、全N等影响土壤肥力的重要指标上均明显高于半裸地,而人工扁袋护岸的土壤养分仅稍高于半裸地,对土壤肥力的改善不是十分显著。综合分析各岸坡土壤特征因子得出,在维持保水保肥、涵养水源等功能方面,效果最好的是植被与天然织物垫护岸,其效果稍好于人工扁袋,相比原生态裸地均有较明显的改善。
2.3 不同护岸模式的生物多样性特征值与土壤特征因子关系的分析
设土壤容重为X1,毛管孔隙度为X2,非毛管孔隙度为X3,全 N 为 X4,全P 为X5,全K 为 X6,速效N为 X7,速效 P为 X8,速效 K 为 X9,pH 值为 X10,有机质为X11,阳离子交换量为 X12,将这12种土壤特征因子与生物多样性的4各指标特征值Margalef丰富度指数(R)、Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)、Pielou均匀度指数(E)进行一般相关关系分析(见表4)。通过相关关系分析,各多样性指标均与土壤容重呈显著性负相关;Margalef丰富度指数、Simpson优势度指数与毛管孔隙度、速效N、CEC呈显著性正相关,与有机质呈极显著性正相关;Shannon-Wiener多样性指数、Pielou均匀度指数与毛管孔隙度呈极显著的正相关,与有机质呈显著性正相关;其中,Shannon-Wiener多样性指数还与速效N呈显著性正相关;各多样性特征值与非毛管孔隙度、全N、全P、全 K、速效P、速效K、pH 的相关关系并不明显。
进一步采用逐步回归法(见表5)分析不同护岸模式的植物群落物种多样性指数与土壤特征因子的关系,并构建多元线性回归模型发现,不同护岸模式的多样性指数与土壤特征因子关系极其显著,尤其是全K、有机质和阳离子交换量CEC三个土壤养分因子均进入了Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数所建立的回归方程中,表明植物多样性对提高土壤养分的作用十分显著。

表4 各岸坡植被多样性指数与土壤特征因子的相关性

表5 各岸坡植被多样性指数与土壤特征因子的逐步线性回归方程
3 结论
(1)对各护岸模式的生物多样性指数特征值进行平行分析,植被护岸与天然材料织物垫护岸的物种多样性最为丰富。乔木层,由于只有植被护岸栽植大量乔木,其余岸坡栽植很少或者无乔木,因此乔木的生物多样性并无代表性。灌木层,物种多样性指数以植被(1.605)和天然织物垫(1.532)护岸最大,远高于原生态半裸地。草本层,各护岸模式的Margalef丰富度指数、Shannon-Wiener多样性指数变化规律基本一致,各新增护岸模式的多样性远比半裸地要好。
(2)纵向比较,除了Pielou均匀度指数外,草本群落层次的各多样性指数值均高于灌木群落层次,对群落样方调查发现,草本层次中,虽然引进的优势种数不及灌木层次引进的物种数丰富,但其数量较多;而在灌木层次中,虽然群落在垂直结构上增加了灌木层,但由于草本植物更容易自然演替,物种数增长更快,且数量远多于乔灌层,在一定程度上掠取了乔灌木的生长空间与养分供给,使下层草本与上层的乔灌木生长相互抑制,使灌木层的总体多样性低于草本群落。
(3)从群落总体分布来看,各新增护岸措施相比为治理前的原生态半裸地多样性明显增加,从草本群落向乔灌草混合群落逐渐演替,群落的垂直结构也发生了明显变化,增加了乔木层,物种的种类和数目明显增多。同时,为了满足更多物种对群落资源的需求,群落中资源在人为辅助的作用下发生了重新分配。
(4)各护岸模式灌木层多样性指数规律性并不十分显著,这是因为示范区内的优势灌木是人为引进并栽植分布的,且逐行分布均匀,导致均匀度指数达到了0.95以上,具有明显的人为干扰痕迹。草本层中多样性指数规律的不确定性除了人为引进的优势种黑麦草、苏丹草外,不同坡位导致杂草生存的外部环境条件不同,尤其是近水区的土壤含水率和营养成分更丰富,而各种杂草对该外部环境条件改变的适应能力却有所差异。
(5)从各护岸模式的土壤特征因子关系来看,各岸坡的土壤特征因子呈显著性差异,其中植被护岸与天然材料织物垫护岸的毛管孔隙度最高,其透水持水能力最强。在保持水土功能方面,植被和天然材料织物垫护岸也是最好的,这两种护岸模式的土壤养分因子极明显的高于半裸地,说明其保持水肥的效果突出。
(6)根据国内外学者研究报告,Gentry[20]认为随着土壤肥沃程度的增加,群落的物种多样性也逐渐增加,王长庭[5]等研究认为土壤容重随着物种多样性的增加而减少。这均与本文的研究结果一致。通过对多样性指数与土壤特征因子进行逐步回归分析,全K、有机质和阳离子交换量CEC均进入了Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数所建立的回归方程中,与各多样性指标的关系最为密切,说明土壤养分对多样性的影响更显著。
[1] 傅伯杰,陈利顶,马克明,等.景观生态学原理及应用[M].北京:科学出版社,2001.
[2] 茹广欣,朱登强,王军辉,等.西藏色季拉山急尖长苞冷杉林地的物种多样性与土壤养分特征[J].河南农业大学 学报 ,2008 ,42(5):511-515.
[3] 王凯博,陈美玲,秦娟,等.子午岭植被自然演替中植物多样性变化及其与土壤理化性质的关系[J].西北植物学 报 ,2007 ,27(10):2089-2096
[4] Ansgar Kahmen,Jorg Perner,Volker Audorff,et al.Effects of plant diversity,community composition and environmental parameterson productivity in montane European grasslands[J].Oecologia,2005,142:606-615.
[5] 王长庭,曹广民,王启兰,等.三江源地区不同建植期人工草地植被特征及其与土壤特征的关系[J].应用生态学报,2007,18(11):2426-2431.
[6] 左小安,赵学勇,赵哈林,等.科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系[J].环境科学,2007,28(5):945-951.
[7] 文海燕,傅华,赵哈林.退化沙质草地植物群落物种多样性与土壤肥力的关系[J].草业科学,2008,25(10):6-9.
[8] 刘克彪.盐渍化沙地土壤旱化过程中植被的变化[J].草业科学,2005,22(10):7-10.
[9] 肖德荣,田昆,张利权.滇西北高原纳帕海湿地植物多样性与土壤肥力的关系[J].生态学报,2008,28(7):3116-3124.
[10] 赵洋毅,王玉杰,王云琦,等.渝北不同模式水源涵养林植物多样性及其与土壤特征的关系[J].生态环境学报,2009,18(6):2260-2266.
[11] 汪冬冬,杨凯,车越,等.河段尺度的上海苏州河河岸带综合评价[J].生态学报,2010,30(13):3501-3510.
[12] 陈小华,李小平.河道生态护坡关键技术及其生态功能[J].生态学报,2007,27(3):1168-1176.
[13] 宋创业,郭柯,刘高焕.浑善达克沙地植物群落物种多样性与土壤因子的关系[J].生态学杂志,2008,27(1):8-13.
[14] 陈圣宾,蒋高明,高吉喜,等.生物多样性监测指标体系构建研究进展[J].生态学报,2008,28(10):5124-5132.
[15] 章家恩.生态学常用实验研究方法与技术[M].北京:化学工业出版社,2006:91-92.
[16] 张金屯.数量生态学[M].北京:科学出版社,2004.
[17] 杨万勤,钟章成,陶建平,等.缙云山森林土壤速效P的分布特征及其与物种多样性的关系研究[J].生态学杂志,2001,20(4):24-27.
[18] 王占军,蒋齐,潘占兵,等.宁夏毛乌素沙地不同密度柠条林对土壤结构及植物群落特征的影响[J].水土保持研究,2005,12(6):123-125.
[19] 李德生,张萍,张水龙,等.黄前库区森林地表径流水移动规律的研究[J].水土保持学报,2004,18(1):78-81.
[20] Gentry A H.Endemism in tropical vs.temperate plant communities in Soule[M].Sunderland,Massachusetts:Conservation Biology.Sinauer Press,1986:153-181.
猜你喜欢
材料与冶金学报(2022年2期)2022-08-10 09:15:38
纺织科学研究(2021年7期)2021-12-02 02:56:27
华南农业大学学报(2021年4期)2021-07-24 06:02:14
纺织科技进展(2021年5期)2021-07-22 08:41:38
干旱地区农业研究(2019年3期)2019-07-18 07:09:40
水利规划与设计(2017年9期)2017-12-20 08:24:57
山东工业技术(2017年21期)2017-11-04 21:21:04
水利技术监督(2016年6期)2017-01-15 14:01:41
水利科技与经济(2016年10期)2016-04-26 08:40:22
中国洗涤用品工业(2015年8期)2015-02-28 19:02:53
