
荒漠区子午沙鼠种群数量动态及其预测
2011-04-25 09:59:38张福顺付和平武晓东张晓东杨泽龙
草业科学 2011年3期
张福顺,付和平,武晓东,袁 帅,张晓东,杨泽龙
(1.内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019; 2.内蒙古自治区气象科学研究所,内蒙古 呼和浩特 010051)
荒漠生态系统极其脆弱,鼠害的发生会加剧其退化。然而,只有当啮齿动物数量超过危害阈值时才形成危害。在危害阈值之内,不仅不会对生态系统造成危害,还有益于维持生态系统的平衡[1-2]。因此,利用啮齿动物多年的种群变动规律进行种群数量预测研究,将啮齿动物种群控制在危害阈值以内,对于荒漠生态系统的平衡与稳定具有重要意义。
国内有关啮齿动物种群动态和预测预报的研究主要集中在草原和农田上,董维惠等对荒漠草原地区长爪沙鼠(Meriones unguiculatus)[3]、黑线仓鼠(Cricetulus barabensis)[4]、五趾跳鼠(Allactaga sibirica)[5]和三趾跳鼠(Dipus sagitta)[6]等进行了较多研究,而关于荒漠地区的研究报道较少。子午沙鼠(M. meridianus)是荒漠区主要优势种之一,许多学者对该鼠种的食性、繁殖、生态习性和种群数量动态等进行了研究[7-13],但是关于该鼠种在荒漠地区的研究鲜见报道。本研究利用多年的野外调查数据,对阿拉善荒漠地区禁牧区和轮牧区子午沙鼠的种群动态进行分析,同时利用线性回归分析进行种群预测研究,旨在为该地区啮齿动物的防治提供指导依据。
1 材料与方法
1.1研究区自然概况 研究区位于内蒙古阿拉善左旗南部的嘉尔格勒赛汗镇,地理坐标为104°10′~105°30′E,37°24′~38°25′N,地处腾格里沙漠东缘。该地区的草地类型是典型的温性荒漠,植被稀疏,结构单调,覆盖度低,一般为1%~20%。植物种类贫乏,主要以旱生、超旱生和盐生的灌木和小半灌木为主。建群植物以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygophyllaceae)为主,其次为蔷薇科(Rosaceae)、柽柳科(Tamaricaceae)。地形起伏不平,丘陵、沙丘与平滩相间。气候为典型的高原大陆性气候,冬季严寒、干燥,夏季酷热,昼夜温差大,极端最低气温-36 ℃,最高气温42 ℃,年平均气温8.3 ℃,无霜期156d。年降水量45~215mm,且降水极不均匀,主要集中在7-8月。年蒸发量3 000~4 700mm。土壤为棕漠土,淋溶作用微弱,土质松散、瘠薄,表土有机质含量1.0%~1.5%,含有较多的可溶性盐。
1.2研究方法 2002年在原生植被一致的基础上,选取两种不同干扰类型样地,分别为:Ⅰ禁牧区,自1997年开始围封禁牧,到本研究开始的2002年已禁牧5年以上,研究期间始终处于禁牧状态;Ⅱ轮牧区,该样地自1995年开始采取围栏轮牧的利用方式,将500hm2的轮牧区划分为4个轮牧小区,放牧300只羊,各小区每月放牧1次。2002-2010年,每年4、7、10月的上旬采用铗日法对各样地的啮齿动物种群数量进行调查。每次取样时,在各样地随机选择4个铗捕样方,样方面积10hm2。每个样方布设5条铗线,每铗线100铗,铗距5m,铗线距50m,共计500铗日。鼠铗为铁制标准板铗,新鲜花生米作诱饵。鼠铗放置24h后收铗,捕到的标本均进行体尺测量、种类及繁殖状况鉴定。以啮齿动物的捕获率表示种群数量的大小。
2 结果与分析
2.1子午沙鼠种群数量的年度动态 2002-2010年,在禁牧和轮牧两种干扰样地中共布放107 870个铗日,捕获鼠5 948只,分属4科11种,其中捕获子午沙鼠2 360只,占总捕获率的39.68%。在禁牧区,子午沙鼠年均捕获率为37.32%,为优势种;而在轮牧区,子午沙鼠的捕获率为33.34%,仅次于小毛足鼠(Phodopus roborovskii),为次优势种。
2008年禁牧区和轮牧区中子午沙鼠的种群数量相差较大,轮牧区的年均捕获率为禁牧区的3.66倍,其他各年份中相差不大(图1)。2002-2007年,禁牧区中子午沙鼠的种群数量高于轮牧区,而2008-2010年情况相反,轮牧区高于禁牧区。轮牧区中,2008年子午沙鼠的捕获率明显高于其他年份,达到13.25%,为最低年份(2006年)的57倍,其他各年份的种群数量均较低,捕获率均未超过3%;而在禁牧区,2002-2010年子午沙鼠的种群数量波动较小,均低于4%。
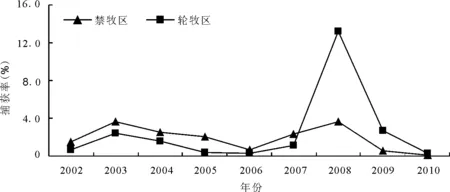
图1 子午沙鼠种群数量的年度变动曲线
2.2子午沙鼠种群数量的季节动态 各年度子午沙鼠种群数量在禁牧区和轮牧区捕获率最高的月份并不相同(图2、3)。禁牧区,2003和2009年的最高月份为7月,2004、2008和2010年最高月份为10月,其余4年最高月份均为4月。轮牧区,2002、2005、2006和2010年各季节捕获率均较低,小于1%。2008年各季节捕获率明显高于其他年份,7月最高,为17.20%,10月次之,为15.45%,4月为7.10%。
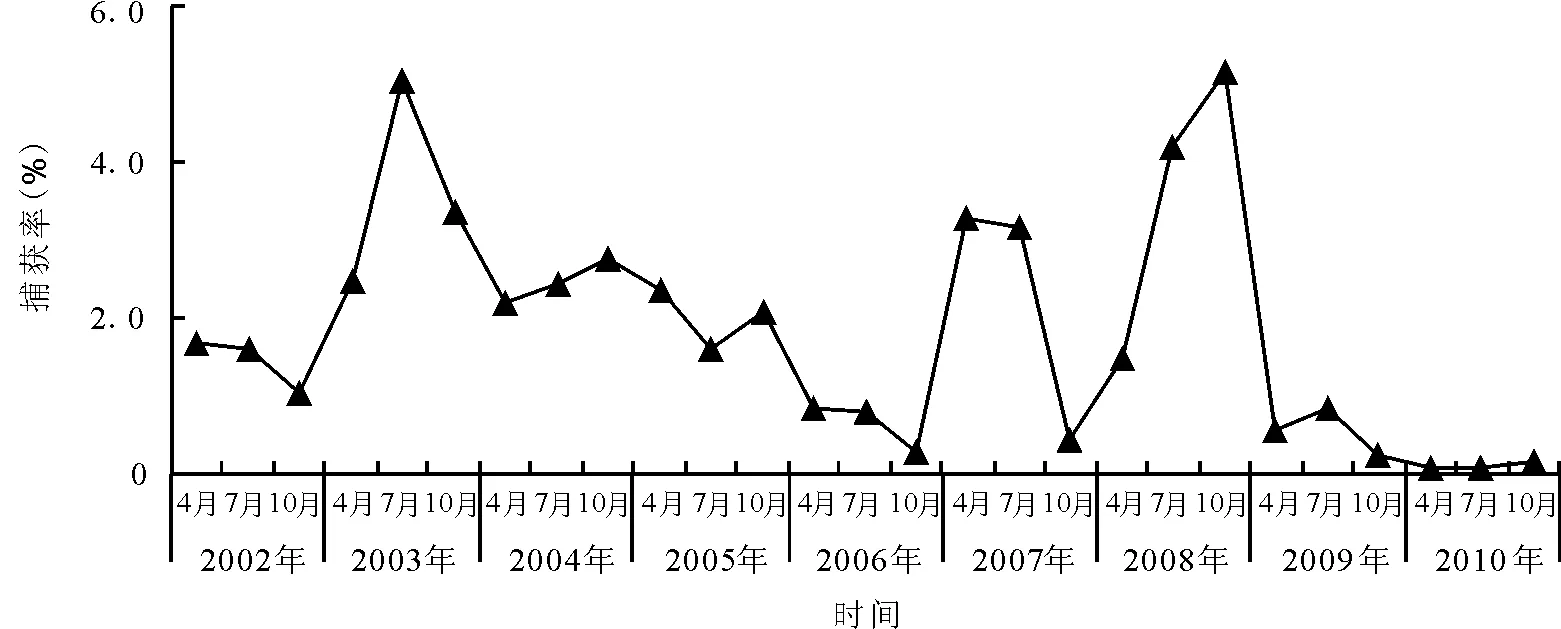
图2 禁牧区子午沙鼠种群数量的季节变动曲线

图3 轮牧区子午沙鼠种群数量的季节变动曲线
对比两种干扰下同一季节的子午沙鼠种群数量(图4),4月禁牧区的种群数量略高于轮牧区,但方差分析结果显示差异不显著(P>0.05),7月和10月轮牧区的种群数量均显著高于禁牧区(P<0.05)。同一干扰下不同季节的种群数量比较得出,禁牧区4、7和10月的种群数量相差不大,捕获率均为2%左右;而轮牧区中7月的捕获率为3.26%,明显高于4和10月。
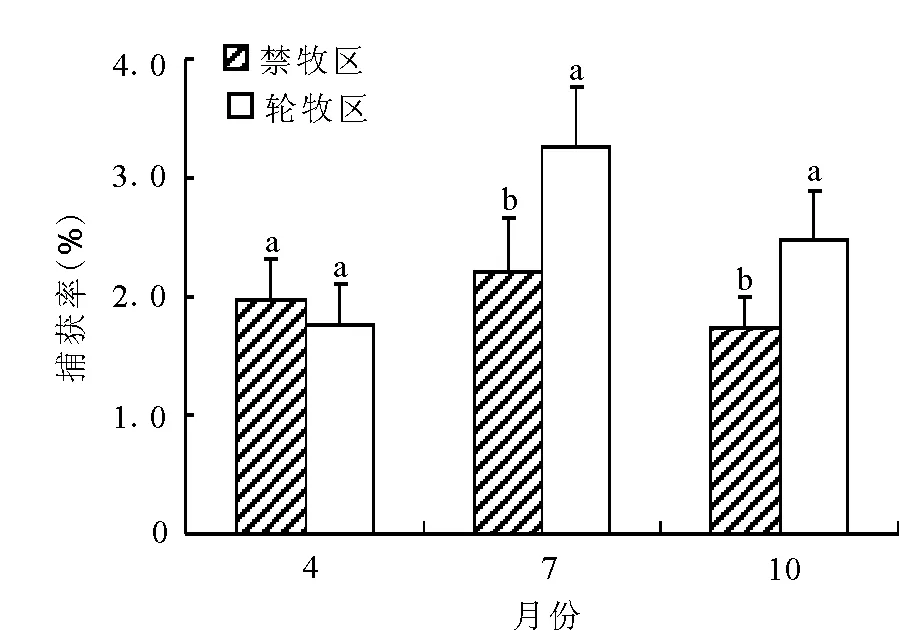
图4 2002-2010年子午沙鼠各季节的平均种群数量
2.3种群数量预测 通过以上分析可以看出,在禁牧区和轮牧区子午沙鼠的种群数量动态具有一定的差异,因此本研究分别对禁牧区和轮牧区子午沙鼠进行种群数量预测。利用各年度4和7月子午沙鼠的捕获率进行一元线性回归分析,对当年7月的捕获率进行预测。禁牧区中4月子午沙鼠的捕获率与7月的捕获率显著正相关(P<0.05),预测模型为y1=1.085 4x1+0.384 9;轮牧区中4与7月的捕获率极显著正相关(P<0.01),预测模型为y2=2.397 3x2-0.974 1(表1)。禁牧区仅2003和2008年的预测值与实测值相差较大(表2),预测准确率为77.78%;轮牧区2003、2004和2007年的预测误差较大,预测准确率为66.67%(表3),但数量高峰年2008年的预测十分准确。

表1 禁牧区和轮牧区的短期预测结果

表2 禁牧区各年4月预测7月捕获率的预测值和实测值
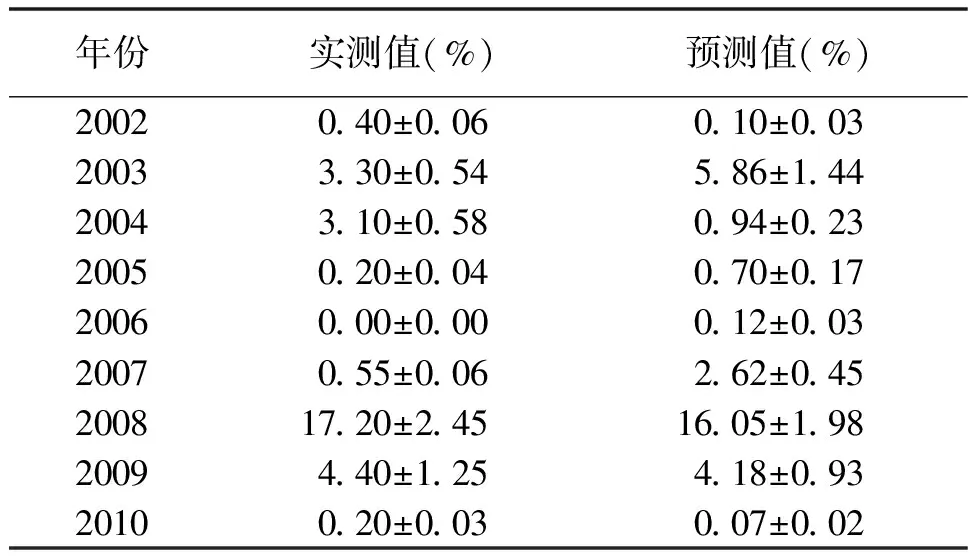
表3 轮牧区各年4月预测7月捕获率的预测值和实测值
3 讨论与小结
种群动态一直是种群生态学研究的核心问题,其中有关小型啮齿动物的研究较多[14]。啮齿动物种群并不是长期维持在一个水平上,而是经常表现为有规律或无规律的波动。种群动态就是研究种群大小或数量在时间上和空间上的变动规律。种群的季节消长比较有规律,而大多数种群的年间变动表现为不规则的波动,具有周期数量变动的种群是有限的[15]。本研究采用9年的野外调查数据,较深入地了解了子午沙鼠在禁牧和轮牧干扰条件下的种群数量变动特征。轮牧区中子午沙鼠的数量仅在2008年出现一个高峰,而禁牧区中9年的种群数量均较低,且未表现出明显的周期性变动,因此关于子午沙鼠种群数量的年间变动在该区域是否具有周期性,还需要进一步的研究。
子午沙鼠种群数量变动在年际和季节上具有差异,其影响因素是多方面的。年际变动的主要差异表现为2008年轮牧区的捕获率明显高于禁牧区。干扰可能是造成差异的主要因素,武晓东与付和平[16]研究表明,啮齿动物对不同干扰的敏感性反应不同,而2008年为子午沙鼠的高峰年,这种敏感性在高峰年中表现的更明显。预测过程中,有些年份的预测准确率较低。例如,禁牧区中,2003和2008年准确率较低,原因可能是2003和2008年为种群变动的两个数量高峰年,4-7月子午沙鼠的捕获率迅速上升,降低了模型预测的准确率。啮齿动物种群波动性较大是造成线性回归预测准确率降低的重要原因,但是禁牧区和轮牧区中子午沙鼠的预测准确率分别为77.78%和66.67%,在实践中具有一定的实用性和可行性。
国内外关于啮齿动物种群数量波动及其影响因素的研究报道较多。赵天飙等[17]研究表明,大沙鼠(Rhombomys opimus)种群数量与降水量有显著的相关关系,且密度的升高和降低总是滞后于降水量。Leirs等[18]用12年数据分析得出,雨季中降水量分布对坦桑尼亚栗鼠(Mastomys natalensis)的种群动态有重要影响,种群数量爆发均出现在雨季早期有丰富降雨的年份中,可见降水量是影响啮齿动物种群动态的因素之一。Murúa等研究表明[19],智利南部米鼠(Oligoryzomys longicaudatus)的种群数量大小受食物、空间和捕食者等因素的限制以及种内竞争调节的影响。侯希贤等[20]发现,小毛足鼠的种群数量变化季节性明显,年间差异较大,其主要影响因素有种群年龄组成中幼年组的百分比、怀孕率、繁殖指数以及其他鼠种的种群数量。Brown和Munger[21]认为,食物资源尤其是一年生植物和种间相互作用对啮齿动物的种群密度具有重要影响。以上影响因素对于本研究中子午沙鼠数量变动的影响是必然的,但本研究认为放牧干扰也是影响种群动态的重要原因。刘伟等[22]的研究也表明,啮齿动物种群密度和放牧强度显著相关,根田鼠(M.oeconomu)和甘肃鼠兔(Ochotona cansus)的种群密度随放牧强度的增加而逐渐降低。总之,影响啮齿动物种群数量变动的因素比较复杂,往往是各种内部因素和外部因素综合作用的结果,而且这些作用可能是非线性的。关于啮齿动物的预测研究,可以首先确定影响啮齿动物种群数量的主要因素,然后利用多元或非线性方法进行预测,将会取得更好的预测效果[23]。
为了恢复草原生态环境,我国于2003年在西部11个省区启动了“退牧还草”工程[24],围栏禁牧和适度轮牧已成为草地利用的主要方式。本研究表明,两种干扰下的子午沙鼠种群动态存在差异,因此分别对两种干扰条件下种群数量进行预测是必要的,这对促进草原生态的良性循环具有实践意义。两种干扰条件下,当年4月和7月的种群数量均显著相关,因此,在实践中草原保护部门可以利用铗日法对4月子午沙鼠的种群数量进行调查,利用4月的种群数量预测7月的种群数量,然后根据预测值采取有效的防治措施,防止啮齿动物爆发所带来的经济和生态损失。
[1] 杨莹博,辛小娟,艾得协措,等.鼢鼠土丘植被恢复演替过程中的物种多样性变化[J].草业学报,2010,19(1):14-20.
[2] 周雪荣,郭正刚,郭兴华.高原鼠兔和高原鼢鼠在高寒草甸中的作用[J].草业科学,2010,27(5):38-44.
[3] 董维惠,侯希贤,杨玉平,等.长爪沙鼠种群数量变动特征的研究[J].中国媒介生物学及控制杂志,2004,15(2):88-91.
[4] 董维惠,侯希贤,林小泉,等.黑线仓鼠种群数量动态预测研究[J].生态学报,1993,13(4):300-305.
[5] 董维惠,侯希贤,杨玉平.内蒙古中西部地区五趾跳鼠种群数量动态研究[J].中国媒介生物学及控制杂志,2006,17(6):444-446.
[6] 董维惠,侯希贤,杨玉平.三趾跳鼠种群数量动态及预测研究[J].中华卫生杀虫药械,2008,14(3):181-184.
[7] 赵肯堂.子午沙鼠的繁殖调查[J].四川动物,1985(2):17-19.
[8] 周延林,王利民,鲍伟东,等.子午沙鼠种群繁殖特征[J].兽类学报,1999,19(1):62-67.
[9] 刘焕金,冯敬义,李承节,等.子午沙鼠生态的调查研究[J].动物学杂志,1984(4):21-25.
[10] 宋恺,刘荣堂.子午沙鼠的生态研究[J].兽类学报,1984,4(4):291-299.
[11] 李枝林,秦长育,韩建芳.子午沙鼠生态学的初步研究[J].兽类学报,1988(1):43-48.
[12] 侯希贤,董维惠,周延林,等.子午沙鼠种群数量动态及预测[J].生态学报,2000,20(4):711-714.
[13] 董维惠,侯希贤,杨玉平.子午沙鼠种群数量动态分析[J].中国媒介生物学及控制杂志,2005,16(1):23-25.
[14] 游群.我国啮齿动物种群生态学研究进展[J].陕西林业科技,2004(1):26-30.
[15] 孙濡泳.动物生态学原理[M].第三版.北京:北京师范大学出版社,2001:200-211.
[16] 武晓东,付和平.人为干扰下荒漠啮齿动物群落格局——变动趋势与敏感性反应[J].生态学报,2006,26(3):849-861.
[17] 赵天飙,杨持,周立志.大沙鼠种群密度与降水量的关系[J].兽类学报,2007,27(2):195-199.
[18]LeirsH,VerhagenR,VerheyenW,et al.ForecastingrodentoutbreaksinAfrica:anecologicalbasisforMastomyscontrolinTanzania[J].JournalofAppliedEcology,1996,33(5):973-943.
[19]MurúaR,GonzálezLA,LimaM.Populationdynamicsofricerats(aHantavirusreservoir)insouthernChile:feedbackstructureandnon-lineareffectsofclimaticoscillations[J].Oikos,2003,102(11):137-145.
[20] 侯希贤,董维惠,杨玉平.鄂尔多斯沙地草场小毛足鼠种群数量动态分析[J].中国媒介生物学及控制杂志,2003,14(3):177-179.
[21]BrownJH,MungerJC.Experimentalmanipulationofadesertrodentcommunity:foodadditionandspeciesremoval[J].Ecology,1985,66(5):1545-1563.
[22] 刘伟,周立,王溪.不同放牧强度对植物及啮齿动物作用的研究[J].生态学报,1999,19(3):376-382.
[23] 王玮,冯琦胜,于惠,等.“3S”技术在草地鼠虫害监测与预测中的应用[J].草业科学,2010,27(3):31-39.
[24] 包利民.我国退牧还草政策研究综述[J].农业经济问题,2006(8):61-65.
猜你喜欢
疯狂英语·新读写(2024年2期)2024-03-20 01:41:16
海外星云(2021年6期)2021-10-14 07:20:46
意林·少年版(2021年13期)2021-08-30 16:26:41
西藏艺术研究(2021年3期)2021-06-02 09:37:16
动漫星空(兴趣百科)(2020年5期)2020-06-10 03:12:02
西藏农业科技(2019年1期)2019-07-25 00:37:08
浙江医学(2018年16期)2018-09-08 05:58:00
消费导刊(2016年7期)2017-08-12 13:23:32
兽医导刊(2016年6期)2016-05-17 03:50:31
实验动物与比较医学(2014年5期)2014-02-28 14:53:08
