
交配次数对云斑天牛雌虫精子消耗量及产卵量的影响
2011-04-04 03:19王保新杨春平
环境昆虫学报 2011年1期
王保新,杨 桦,2,林 森,杨 伟,杨春平
(1.四川农业大学森林保护重点实验室,四川 雅安 625014;2.贵州大学昆虫研究所,贵州山地农业病虫害重点实验室,贵阳 550025)
交配是昆虫选择最佳配偶以及产生后代的最重要最基本的过程,也是具有重要进化意义的过程。而多次交配 (Multiple mating)是昆虫繁衍后代所采取的一种典型的交配策略 (刘兴平等,2008)。雌虫通过与多个雄虫发生交配行为以补充精子,确保受精的成功而增加雌虫产生更多后代的可能性 (Pai et al.,2005)。然而,多次交配对不同种类昆虫精子消耗量、产卵量的影响各不相同 (Sakubai,1998)。杨洪等 (2006)研究表明,交配一次的松墨天牛Monochamus alternatus雌虫受精囊内精子数量因不断消耗,在交配16 d后只有5万以下,多次交配的松墨天牛雌虫受精囊内的精子数量长期保持在12万个左右从而使产卵量显著大于单次交配的产卵量。新西兰麦长蝽Nysius huttoni的多次交配行为没有明显地影响到雌虫的产卵量 (Wang and Davis,2006)。而小菜蛾Plutella xylostella雌虫的产卵量随交配次数的增加反而减少(Wang et al.,2005)。
云斑天牛Batocera horsfieldi(Hope)隶属于鞘翅目Coleoptera天牛科Cerambycidae,又名云斑白条天牛,危害杨树Populus spp.、核桃Juglans regia等多种树木,是我国重要林木蛀干害虫 (李娟等,2008)。有关云斑天牛的多次交配和精子消耗的研究未见报道,本研究以云斑天牛为对象,研究多次交配对云斑天牛受精囊内精子数量消耗、产卵量的影响,以期为天牛多次交配机理和进化机制的深入研究奠定基础。
1 材料与方法
1.1 供试虫源
2010年5月,于四川省德阳市罗江县采集刚羽化未交配的杨树云斑天牛成虫放入60 cm ×60 cm×60 cm的不锈钢网养虫笼内。于室温下用200 mL三角瓶水培法国冬青枝条饲养成虫备用,每3 d更换1次冬青枝条,用直径4~5 cm,长约20 cm新鲜杨树木段作为产卵诱木,3 d更换1次。已交配雄虫腹部绒毛磨损,形成明显的交配斑;已交配雌虫为背部绒毛磨损,形成明显交配斑(嵇保中等,1996)。
1.2 雌性生殖系统及精子形态观察
取羽化后10 d的3雌3雄云斑天牛成虫,剪下雌虫腹部,置于30%NaOH溶液中煮10 min,取出生殖系统,用Sz66体视显微镜观察并拍照。同时在载玻片上解剖雄虫,取出睾丸,用解剖针刺破睾丸后,用0.9%生理盐水稀释,观察精子的形态并拍照。
1.3 单次交配后精子在雌性生殖系统内的分布动态观察
分别于雌虫单次交配 (单次交配指成功交配一次,包括交配前求偶、插入、配后保护等完整阶段) 后10 min、1 h、2 h、4 h、8 h、16 h、32 h解剖生殖系统,取下生殖腔、交配-受精囊,并将其分别置于载波片上,用解剖针挑破后用0.9%生理盐水冲洗3次,把冲洗液全部转入1.5 mL离心管内,定容到0.5 mL,摇匀后取10 μL混匀液滴于血球计数板上,观察统计精子的数量。每份混匀液统计3次,取平均值。每个观察时间雌虫样本数为3头。
1.4 单次和多次交配雌性生殖系统内精子数量的观察
雌虫单次和多次交配 (配对饲养)后,分别于不同时间 (1 d、2 d、4 d、8 d、16 d)观察统计交配-受精囊和生殖腔内精子的数量。每个观察时间雌虫取样数为3头。
1.5 单次和多次交配雌虫产卵量变化的观察
单次交配:选同期羽化后10 d的云斑天牛,以1♀∶1♂配对,观察交配情况。取30头成功交配一次的雌虫单独饲养,待雌虫开始产卵后每天记录每头雌虫的产卵量。
多次交配:参照张世渊和路高 (2001)、杨洪等 (2006)对松褐天牛的研究,选同期羽化后10 d的云斑天牛,以1♀∶1♂配对饲养,待雌虫开始产卵后,记录每头雌虫的产卵量。共30个重复。
1.6 数据统计分析
试验所得数据用SPSS 13.0进行统计分析,采用Microsoft Office Excel 2003绘图,单次交配和多次交配雌虫产卵量的比较采用独立样本t检验比较。
2 结果与分析
2.1 雌性生殖系统及精子形态
云斑天牛雌性生殖器由一对卵巢、一对侧输卵管、中输卵管、阴道、交配囊、受精囊、受精囊腺、生殖腔、骨针和产卵器 (图1)。
交配囊圆柱形,基部经交配囊柄与阴道相连。受精囊管侧生于交配囊上,受精囊管上连有细长管状的受精囊腺和“C”形的受精囊。受精囊外壁骨化,顶部指状膨大。受精囊与受精囊管外部有一共同膜包被,使受精囊外观呈肾形 (嵇保中和钱范俊,1995)。

图1 云斑天牛雌性生殖系统Fig.1 Reproductive systems of female B.horsfieldi
云斑天牛精子形状见图2,精子为鞭毛型,丝状,长0.5 mm左右,头部不明显,颜色较深可分辨。
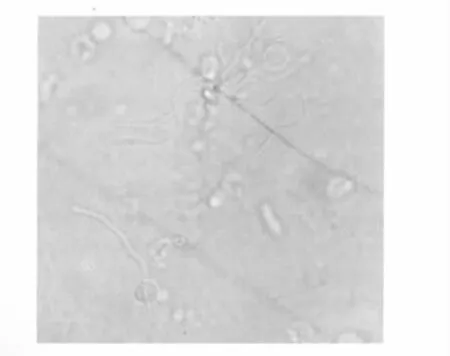
图2 云斑天牛精子形态Fig.2 Spermatozoon morphology of B.horsfieldi
2.2 单次交配精子在雌性生殖系统内的分布
单次交配雌性生殖系统内精子分布见图3。云斑天牛完整交配一次,雄虫输入到雌虫体内的精子大约100万个。这些精子由雄虫输送到雌虫交配囊,然后再由交配囊进入受精囊。16 h后在交配-受精腔内的精子数量基本不变,在受精囊腺分泌物的滋养下可以长期存活至雌虫死亡。在生殖腔内的精子随时间的延长变化不大,基本维持在8万左右。

图3 云斑天牛精子在雌性生殖系统内的分布(平均值±标准误)Fig.3 Distribution of spermatozoa in reproductive system of B.horsfieldi(Mean±SE)
2.3 单次和多次交配精子在雌虫生殖系统内的分布
成功完成一次交配后,云斑天牛雌虫交配-受精腔和生殖腔内的精子数量随着天数的增加而下降,尤其是精子在交配-受精腔内的数量消耗与时间呈线性关系,为y=594780-91810x,(R2=0.9583)。而多次交配的雌虫交配-受精腔内的精子数量保持在80万左右,生殖腔内的精子保持在8万左右,且不随时间的增加而降低 (图4)。说明多次交配对雌虫生殖系统内精子数量有明显补充。

图4 单次交配与多次交配在雌性生殖系统内的分布Fig.4 Quantitative variation of spermatozoa in reproductive system of B.horsfieldi after multiple and single mating(Mean±SE).A,交配 -受精腔;B,生殖腔 (平均值±标准误)A,Bursa coplatrix-Spermatheca;B,Genital cavity.
2.4 单次和多次交配雌虫产卵量的变化
云斑天牛多次交配的雌虫周平均产卵量在8粒左右,产卵历期为30 d,而单次交配的雌虫周平均产卵量在7粒左右,产卵历期为26 d。多次交配的云斑天牛雌虫在周平均产卵量、总产卵量和产卵历期上都显著高于单次交配的云斑天牛雌虫 (P<0.05),多次交配的雌虫寿命与单次交配的雌虫寿命相比较,差异不显著 (P>0.05)(表1)。

表1 云斑天牛单次和多次交配产卵量、产卵历期和寿命的差异 (平均值±标准误)Table1 Variation of fecundity,period of oviposition and longevity between multiple and single females of B.horsfieldi(Mean±SE)
3 结论与讨论
多次交配包括雌虫与同一个雄虫多次交配和与不同的雄虫多次交配。在自然界中多次交配行为广泛存在于昆虫 (Parker,1970;Choe,1995;Imhof et al.,1998)、鸟类(Birkhead et al.,1999)、甚至哺乳类动物中 (Gomendio and Roldan,1991)。本文通过对雌虫单次交配和多次交配的研究发现,云斑天牛雌虫多次交配对雌虫受精囊内精子的贮藏量有明显的补充,这同杨洪等 (2006)对松褐天牛雌虫多次交配的研究一致。
雌虫多次交配的原因主要有提高自身适合度、增加后代遗传多样性和增进生殖成功率等 (Arnqvist and Nilsson,2000)。多次交配对不同种类昆虫的雌虫精子数量消耗、产卵量、孵化率和寿命之间的影响各不相同 (Sakubai,1998)。缺翅目昆虫Zorotypus barberi雌性与同一雄性交配多次时,产卵量比交配一次或两次时多,因而认为雌性与同一雄性多次交配时,是为了获得更多的精子,但与不同雄性多次交配时,可能是为了稀释劣质雄性的精子 (Choe,1995)。鳞翅目昆虫Plodia interpunctella雌性的生殖力和寿命并没有因多次交配种得到精子以外的营养补充而得以提高 (Cook,1999)。多次交配的云斑天牛雌虫交配-受精腔内的精子数量保持在80万左右,其生殖力也显著高于单次交配。云斑天牛雌虫是否存在不同的产卵模式,单次交配的雌虫精子在交配-受精腔内的消耗是否与生殖力的下降有关需要更进一步的研究。
多次交配对雌虫寿命的影响原因极为复杂,包括交配行为的物理损伤和精液附带的其它物质等 (Eberhard,1996)。本研究发现云斑天牛雌虫多次交配后寿命短于单次交配,这与“除了交配饲喂类型外,多数昆虫随着交配次数的增加,雌虫寿命缩短”(Arnqvist and Nilsson,2000)的结论一致。
云斑天牛受精囊极小,外壁骨化,而且紧挨交配囊,不容易将两者分开。因此本试验在统计精子数量时,是计算的交配囊和受精囊内共有的精子数量,没有分别统计交配囊和受精囊内精子,是本试验的不足之处,也是以后进一步研究的重点。
References)
Liu XP,Peng JH,He HM,Xiao HJ,Xue FS,2008.The effect of multiple mating on mitness in insects.Acta Agriculturae Universitatis Jiangxiensis,30(4):592-600.[刘兴平,彭接辉,何海敏,肖海军,薛芳森,2008.多次交配对昆虫适应性的影响.江西农业大学学报,30(4):592-600]
Pai A,Bennett L,Yan G,2005.Female multiple mating for fertility assurance in red flour beetles(Tribolium castaneum).Can.J.Zool.,83(7):913-919.
Sakubai T,1998.Variation in time to sperm depletion and oviposition pattern in female of Riptortus clavatus(Heteroptera:Alydidae).Ann.Entomol.Soc.Am.,91:737-740.
Yang H,Wang JJ,Zhao ZM,Yang DM,Zhang H,2006.Effects of multiple mating on quantitative depletion of spermatozoa,fecundity and hatchability.Zoological Research,27(3):286-290.[杨洪,王进军,赵志模,杨德,张宏,2006.多次交配对松褐天牛精子数量消耗、产卵量和孵化率的影响.动物学研究,27(3):286 -290]
Wang Q,Davis LK,2006.Females remate for sperm replenishment in a seed bug:Evidence from offsp ring viability.J.Insect Behav.,19(3):337-346.
Wang XP,Fang YL,Zhang ZN,2005.Effect of male and female multiple mating on the fecundity,fertility and longevity of diamondback moth,Plutella xylostella L..J.Appl.Entomol.,129(1):3942.
Li J,Wang MQ,Zhang ZC,Chen JY,Zhang GA,2008.Behavioral response of Batocera horsfieldi adults to plant volatiles.Scientia Silvae Sinicae,44(6):168-170.[李娟,王满囷,张志春,张志春,陈京元,张国安,2008.云斑天牛成虫对植物气味的行为反应.林业科学,44(6):168-170]
Ji BZ,Qian FJ,Yan AJ,1996.Improve method of research to Batocera horsfieldi.For.Pest Dis.,(1):45-46.[嵇保中,钱范俊,严敖金,1996.云斑天牛研究方法的改进.森林病虫通讯,(1):45 -46]
Zhang SY,Lu G,2001.The mating and oviposition behavior,egg stage and hatchability of Monochamus alternatus.For.Pest Dis.,1:34-36.[张世渊,陆高,2001.松褐天牛交尾产卵行为和卵期孵化率的测定.森林病虫通讯,1:34-36]
Ji BZ,Qian FJ,1995.Study on reproductive systems of Batocera horsfieldi.Journal of Nanjing Forestry University,19(4):14-19.[嵇保中,钱范俊,1995.云斑天牛生殖系统的研究.南京林业大学学报,19(4):14-19]
Parker GA,1970.Sperm competition and its evolutionary consequences in the insects.Biol.Rev.,45:525-567.
Choe JC,1995.Courtship feeding and repeated mating in Zorotypus barberi(Insecta:Zoraptera). Anim. Behav.,49(6):1511-1520.
Imbof M,Harr B,Brem G,Schlotterter C,1998.Multiple mating in wild Drasophila molamogaster revisited by microsatellite analysis.Mol.Ecol.,7(7):915-917.
Birkhead TR,Martinez JG,Burke T,Froman DP,1999.Sperm mobility detemines the outcome of sperm competition in the domestic fowl.Proc.R.Soc.Lond.B,266:1759-1764.
Gomendio DT,Roldan ER,1991.Sperm competition influences sperm size in mammals.Proc.R.Soc.Lond.B,243(1308):181-185.
Arnqvist G,Nilsson T,2000.The evolution of polyandry:multiple mating and female fitness in insects.Anim.Behav.,60:145-164.
Cook PA,1999.Sperm numbers and female fertility in the moth Plodia interpunctella(Huebner) (Lepidoptera:Pyralidae).J.Insects Behav.,12(6):767-780.
Eberhard WG,1996.Female Control:Sexual Selection by Cryptic Female Choice.Princetom,New Jersey:Princeton University Press.
猜你喜欢
热带生物学报(2022年6期)2022-11-29
甘肃农业科技(2021年6期)2021-07-25
环境昆虫学报(2020年5期)2020-11-11
科学导报(2019年35期)2019-09-03
中国生殖健康(2019年2期)2019-08-23
西北农林科技大学学报(自然科学版)(2018年5期)2018-05-24
中国当代医药(2015年22期)2015-03-01
江苏农业科学(2014年3期)2014-07-16
河南医学研究(2014年5期)2014-02-27
中国医药指南(2010年34期)2010-09-19
