
小尺度条件下退化绢蒿种群幼苗更新时空格局
2011-03-27 06:52:40鲁为华朱进忠靳瑰丽
草业学报 2011年5期
鲁为华,朱进忠,靳瑰丽
(1.石河子大学动物科技学院,新疆 石河子832003;2.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐830052;3.新疆草地资源与生态重点实验室,新疆 乌鲁木齐830052)
幼苗的出苗格局往往和种子的形成以及传播过程密切相关[1],种子落粒过程要受到母株状态的影响[2-4]。有些植物种子只能在短距离内传播甚至传播距离为零。有学者认为短距离的种子传播对于植物种群拓展而言是不成功的[5]。但实际上短距离扩散有利于在环境均一的条件下种群内斑块的形成,同时也有利于竞争种在异质性环境中共存[6-9]。
种子传播严格来讲是属于种群水平上发生的行为,单株传播后形成的格局有可能由于母株个体数量太少而使得随机效应被无形中放大。因此如何用同一种分析方法将多个个体纳入到分析样本中而实现对种子扩散过程的确切描述是一个现实问题。Thorsten Wiegand研究组利用“多样方图像整合技术”(combining the data from individual mapped replicate plots into mean,以下简称 CIM)成功实现了多个小尺度样方的点格局分析,之后并以这种方法进行了大量的研究工作[10]。
近年来,众多学者对绢蒿(Seriphidium)荒漠进行了大量的研究工作,包括绢蒿荒漠的植被动态变化[11]、绢蒿种子结构特征及萌发特性[12,13]、退化过程中土壤和植被的变化规律[14,15]以及绢蒿生物学和生理学特性[16,17]、幼苗的群体变化规律[18]、成株的分布特征[19]等。但对绢蒿种子传播后形成的幼苗空间分布格局未进行充分研究。因此,对绢蒿种群的种子传播后发生的幼苗格局进行小尺度分析,揭示种子短距离扩散后形成的幼苗在不同尺度上的发生格局,期望从小尺度的空间分布格局中认识到绢蒿种群的某些生态学过程,为该类草地的合理利用和生态恢复提供理论依据。
1 材料与方法
1.1 试验方法
首先在研究区由北向南设置3条样线,统计包括绢蒿在内的植物种类、密度、生物量、盖度等指标,利用聚类分析方法将整个地段划分为重度退化(SD,serious degradation)和极度退化(HD,heavy degradation)2个梯度[19],重度退化区主要以叉毛蓬(Petrosimonia sibirica)为主,植被密度较大。极度退化区域主要以叉毛蓬为主,伴生有骆驼蓬(Peganum)等植物,植被稀疏,并有大量的裸地。两区域内绢蒿个体分布稀疏,可完全消除邻接干扰效应。在2008年整个植物群落的生长旺季,对重度和极度退化2个样地中大小级别相似的绢蒿成年植株和其周围发生的幼苗坐标点进行详细记录,每个样地取10个单株,2样地共计20个单株。记录时以单个母株为中心点,布置1m×1m的小样方,尽量将所有幼苗控制在样方范围内。以样方正南正北方向,1m×1m样方的左下角为坐标位置的参考点,按地上绢蒿幼苗的个体记录,只要有绢蒿幼苗个体出现,就把该点坐标位置赋予绢蒿,并对每一绢蒿幼苗坐标位置进行标签标注。
在对绢蒿幼苗定位的基础上,按月统计样方内幼苗的发生数量以及死亡状况,统计其数量的动态变化。
1.2 数据处理
采用生存分析的Kaplan-Meier来统计幼苗存活率大小,并利用Log-Rank秩相关以及Breslow检验来比较不同退化梯度下的幼苗存活能力差异[20];利用Programita 2006软件的O-ring函数进行点格局分析,同时结合软件包内的CIM技术将若干单株幼苗的点格局分析结果通过加权平均,将不同单株幼苗的分布状况反映在相同尺度上[21]。该方法中,双变量O-ring 统计的O12(r)可以用公式(1)计算,该公式通用于单变量和双变量O-ring统计过程。
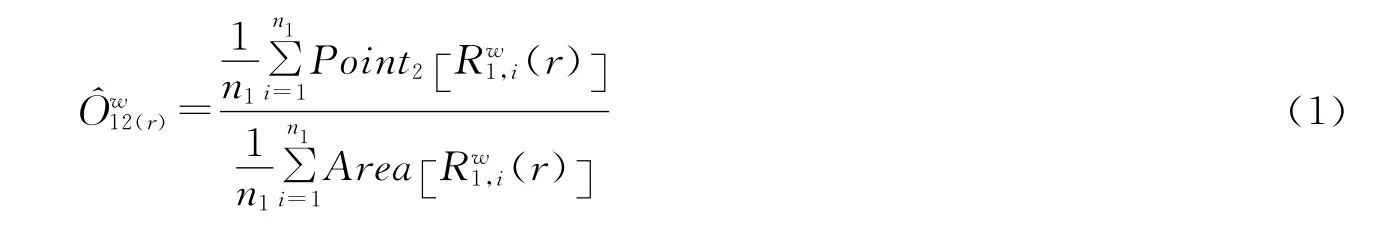
式中,n1为格局1(双变量统计中的对象1)的点的数目;Rw1,i(r)表示以格局1中第i个点为圆心、半径为r、宽为w的圆环;Point2[X]计算了区域X中格局2(双变量统计中的对象2)的点的数目;Area[X]是区域X的面积。
为了将多个重复且进行相同处理的样本等权纳入到计算公式,根据Thorsten Wiegand的意见,在计算多个重复小样方时,可以将公式(1)拓展为公式(2)。
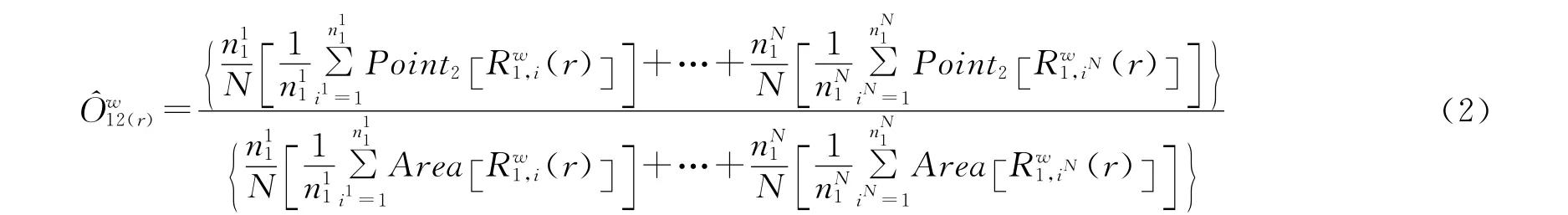
公式(2)中,ij为第j个重复样方的第i个点,n1j为格局1和重复j的点数目,N=∑jn1j为所有重复样方中格局1的点总数目,而公式(2)又可进一步简化为公式(3)。
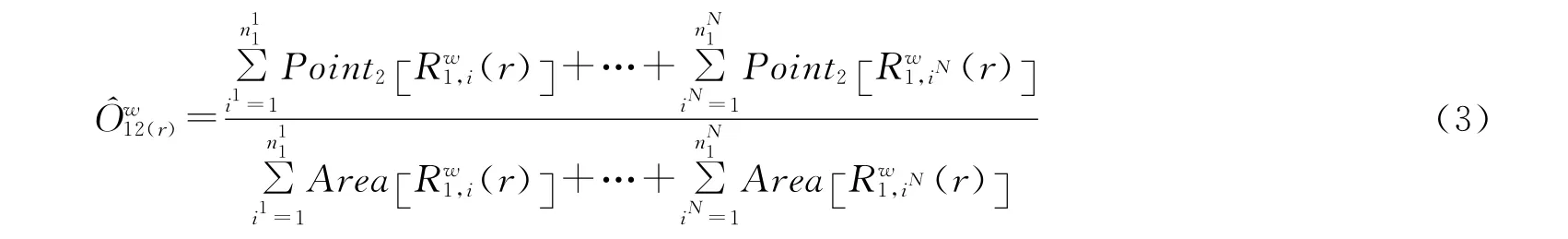
为检验O12(r)偏离零值的显著程度,可以用Monte Carlo随机模拟方法拟合它们95%的置信区间。用r作为横坐标,上下包迹线作为纵坐标绘图,置信区间一目了然。用种群实际分布数据(点图)计算得到不同尺度下的O12(r)值若在包迹线以内,则符合随机分布;若在包迹线以上,则为聚集分布,如在包迹线以下则为均匀分布。
2 结果与分析
2.1 两种退化梯度下单株产生幼苗数量动态变化及生存率分析
从2008年4月份开始,按月对所选定的植株以母株为中心定点观测幼苗数量变化状况,统计幼苗死亡数量和新出现的幼苗数量,并采用Log-Rank和Breslow检验来分析不同退化梯度下幼苗的生存力。
2种退化梯度下的幼苗数量存在差异(图1),但其数量随时间变化动态趋势相似,在4月份有1个萌发高峰,完成整个萌发量的90%以上,5月份在数量上变化不大,但略微增长,之后由于大规模死亡而数量迅速下降。因此,不论在哪种条件下都呈现出“爆发性出苗”和“骤然性死亡”的特点。
为进一步比较2种退化梯度下幼苗的生存率大小,采用医学统计中常用的生存率分析来比较样本之间的生存率差异,2种退化梯度下的Kaplan-Meier生存率曲线自7月份开始出现差别(图2),Log-Rank检验结果中,卡方值=0.275 5,P=0.599 6>0.05,因此2种退化梯度下生存曲线不存在差别。同时,因为重度退化样本观察值n=47,小于期望值E=45.289 98,因此可以认为重度退化条件下幼苗死亡率稍高。进一步以观察个体数量为权重进行Breslow检验结果,卡方值=0.0 0 3 2,P=0.954 7,说明在考虑观察对象数量后其生存曲线也不存在显著差异。说明在小尺度下由于单株对幼苗发生和存活提供了同质的庇护条件,造成二者生存率不存在显著差异。

图1 两种退化梯度下单株幼苗数量动态变化及幼苗死亡数量Fig.1 Quantity of survival seedling,mortality of seedling under two gradation degradation
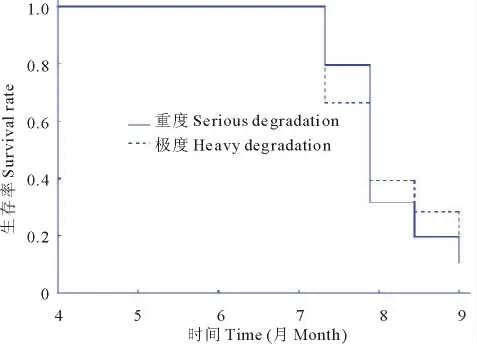
图2 两种退化梯度下幼苗生存率曲线及Breslow检验结果Fig.2 Kaplan-Meier survival rate curve,Log-Rank& Breslow test results in SD and HD
2.2 幼苗空间分布格局分析
将幼苗的点坐标数据转换为以母株为圆心,以最大传播距离为半径的点数据,以探讨幼苗在母株周围的分布状态,并绘制雷达点分布图,由于个体数量限制,这里在2个退化梯度中分别只选取3个较典型植株进行绘图比较,具体结果见图3。
2个退化梯度内的3个单株5月所产生的幼苗数量是比较可观的(图3),但随着时间进程(5-9月)其幼苗数量急剧下降,且存活的幼苗一般都距离母株很接近,重度退化梯度下,5月份幼苗与母株最远距离可以达到近60cm左右,7月份则降至45cm左右,到9月份则缩短至25cm左右。极度退化条件下幼苗的分布也有类似的趋势。
进一步利用Programita 2006软件的O-ring函数和CIM技术对幼苗在不同尺度上的分布状态进行分析(图4)。重度退化条件下,5月份幼苗密度较大,因此在小尺度上聚集强度较大,其密度函数最大值出现在半径为6 cm的圆环上,其值为0.022 2,之后呈直线下降,但在半径小于35cm之前的一系列圆环上,仍然呈现聚集分布,在到达40cm尺度之后转入随机或者均匀分布。到7月份,幼苗数量由于环境筛选而数量减少,因此聚集强度与5月相比显著下降,最大聚集强度出现在半径为6cm的圆环上,其函数最大值为0.016 67,并且偏离聚集分布的尺度提前至33cm;而到9月份,幼苗数量进一步减少,其格局分布虽然是在小尺度上呈聚集分布,但是曲线出现明显的波谷,并且聚集强度总体上进一步减小,其密度函数最大值出现在半径为7cm的圆环上,其值为0.013 02,之后在22cm尺度上转入随机分布。
极度退化条件下,由于幼苗数量随着退化程度增加其数量较少,因此,5月份聚集强度与重度退化比较显著降低,最大聚集强度出现在半径为4cm的圆环上,其值为0.015 3,在到达28cm时,转入随机分布。到7月份,小尺度内聚集强度增加,其O(r)最大值仍然在4cm的圆环上,其值为0.019 1,在25cm时则转入随机分布,37 cm时则转入均匀分布。9月份,曲线整体趋势未发生明显变化,但在4cm圆环上其聚集强度与7月相比略微减小,其O(r)值为0.019 0,在19cm尺度上则转入随机分布。从O(r)值变化曲线可以看出,绢蒿幼苗种子传播距离分布在5~30cm内,其种子集中落在5~10cm的尺度内。当然,不同形体大小的植株种子落粒距离可能会有所不同,但本试验中所选取的植株种子基本在此范围内。随时间推移,母株周围的种子有可能因为其他因素被传播到更远的距离,其潜在生态位空间有可能会加大。
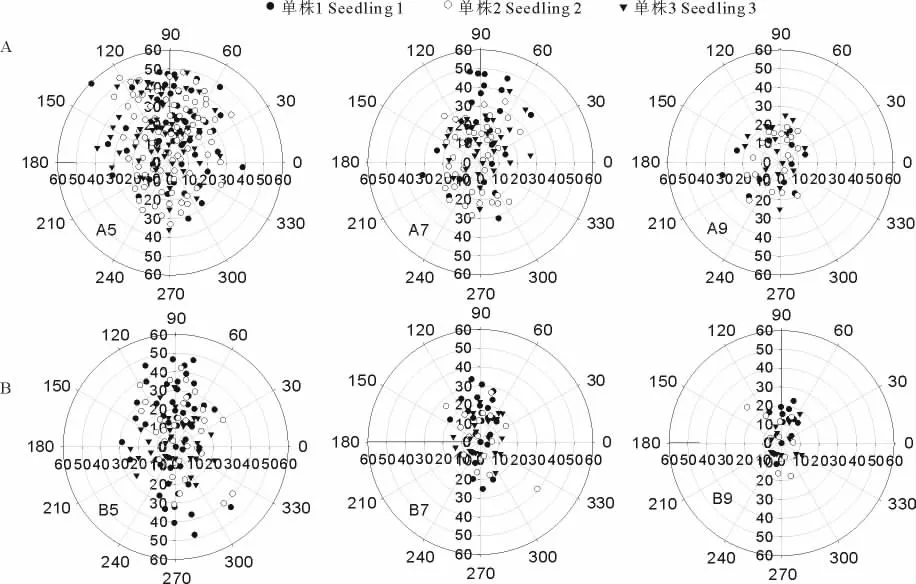
图3 重度(A)和极度(B)退化3个单株产生幼苗5、7、9月的整合点分布图(坐标单位:cm)Fig.3 CIM map of three maternal Seriphidiumseedling in three mounth(Unit:cm)

图4 重度和极度退化条件下3个时间点3个单株幼苗经CIM整合后的点格局分布图Fig.4 Combined spatial patterns of emergence seedling on three maternal piant by CIM in temporal sequence on SD and HD
在2种退化条件下,各时间点上幼苗在空间上的分布格局变化趋势相似,但是经过夏季干旱及高温等环境因子的强烈筛选之后,距离母株较远的幼苗大部分已经死亡,因此在9月份的分布格局与初始格局已经有了明显不同,其有效传播距离尺度缩小,在重度退化区域已经缩小到25cm左右,极度退化区域缩小至20cm左右。从时空尺度来看,绢蒿幼苗对母株具有很强的依赖性,其格局变化总是朝有利于自身生存的方向发展。
3 结论与讨论
绢蒿幼苗在较早时间萌发是一种对中亚荒漠干旱气候的适应性对策,一般在4-5月份土壤中的种子就基本全部萌发,尽早萌发无形中增加了幼苗适合度,因为这种对策使得幼苗个体尽早获得有限的资源并因此而增加其生存的几率并对之后的生长、繁殖等生活史的一系列过程产生积极影响[22,23]。从其生存率来看,大量的幼苗集中萌发为环境筛选提供了足够数量的个体,使得最终还有一部分幼苗存活下来,这将对种群的更新和繁衍提供足够的物质基础。
绢蒿种子落粒对母株具有强烈的依赖性,通过多尺度变换的点格局分析发现,在不同尺度上,幼苗的分布格局是出现变化的,在小尺度上,总是呈现出强烈的聚集分布,而随着尺度增加,其聚集强度逐渐减小,随着时间变化,聚集强度也在发生变化,也就是说,种子向幼苗以及成株转变过程中在时间尺度上面临着强大的环境选择压力,其分布格局总是倾向于使幼苗建成成功率增加的方向转换。
[1]Walker L R,del Moral R,Primary Succession and Ecosystem Rehabilitation[M].Cambridge,UK:Cambridge University Press,2003:442.
[2]Harper J L.The role of predation in vegetational diversity[J].Brookhaven Symposia in Biology,1969,22:48-62.
[3]Crawley.Plant-herbivore dynamics[A].In:Crawley M.Plant Ecology[M].Oxford:Blackwell Science,1997:401-474.
[4]Kitajima K,Fenner M.The Ecology of Regeneration in Plant Communities(2nd edition)[C].Wallingford:CAB International Publishing,2000:331-360.
[5]Cousens R D,Wiegand T.Small-scale spatial structure within patterns of seed dispersal[J].Oecologia,2008,158(3):437-448.
[6]Snyder R E,Chesson P.Local dispersal can facilitate coexistence in the presence of permanent spatial heterogeneity[J].Ecology Letters,2003,6:301-309.
[7]Murrell D J,Law R.Heteromyopia and the spatial coexistence of similar competitors[J].Ecology Letters,2003,6:48-59.
[8]Green D G.Connectivity and complexity in ecological systems[J].Pacific Conservation Biology,1994,1:194-200.
[9]刘钟龄,王炜,梁存柱,等.内蒙古草原植被在持续牧压下退化演替的模式与诊断[J].草地学报,1998,6(4):244-251.
[10]De Luis Martín,Ravents J,Wiegand T.Temporal and spatial differentiation in seedling emergence may promote species coexistence in Mediterranean fire-prone ecosystems[J].Ecography,2008,31:620-629.
[11]刘长娥,许鹏,安沙舟.土质温性荒漠草地植物生育节律分析[J].新疆农业科学,2006,43(1):11-13.
[12]张连义.伊犁绢蒿种子天然种衣吸水保水性能研究[J].植物学报,1998,8:775-777.
[13]孙会忠,宋月芹,贺学礼.绢蒿属8种植物果实微形态特征及其分类学意义[J].武汉植物学研究,2007,25(5):440-444.
[14]顾爱星,范燕敏,武红旗,等.天山北坡退化草地土壤环境与微生物数量的关系[J].草业学报,2010,19(2):116-123.
[15]刘洪来,朱进忠,靳瑰丽.伊犁绢蒿荒漠不同退化阶段草地植物多样性分析[J].新疆农业科学,2007,44(5):632-636.
[16]孙宗玖,安沙舟,许鹏.伊犁绢蒿构件动态变化研究[J].草地学报,2007,15(5):454-459.
[17]孙宗玖,安沙舟,许鹏.伊犁绢蒿体内可塑性营养物质动态变化及分配特征的研究[J].草业学报,2008,17(2):151-156.
[18]鲁为华,朱进忠,王东江,等.天山北坡围栏封育条件下伊犁绢蒿幼苗分布格局及数量动态变化规律研究[J].草业学报,2009,18(4):17-26.
[19]鲁为华,朱进忠,王东江,等.天山北坡两种退化梯度下伊犁绢蒿种群空间分布的点格局分析[J].草业学报,2009,18(5):142-149.
[20]Kaplan E L,Meier P.Nonparametric estimation from incomplete observations[J].Journal of the American Statistical Association,1958,53:457-481.
[21]Wiegand T,Moloney K.Rings,circles and null models for point pattern analysis in ecology[J].Oikos,2004,104:209-229.
[22]Pausas J G.Plant functional traits in relation to fire in crown-fire ecosystems[J].Ecology,2004,85:1085-1100.
[23]Bolker B M,Pacala S W.Spatial moment equations for plant competition:understanding spatial strategies and the advantages of short dispersal[J].American Naturalist,1999,153:575-602.
猜你喜欢
安徽农学通报(2022年6期)2022-04-07 21:30:29
今日农业(2019年13期)2019-08-12 07:59:02
西藏农业科技(2019年2期)2019-07-10 09:23:12
人民调解(2019年5期)2019-03-17 06:55:16
畜牧与饲料科学(2018年3期)2018-05-08 02:48:35
湖南林业科技(2017年1期)2017-02-06 05:28:55
现代园艺·综合版(2016年17期)2016-10-17 04:23:16
果树实用技术与信息(2016年8期)2016-09-21 03:25:35
小说月刊(2015年12期)2015-04-23 08:51:10
湖南农业科学(2014年22期)2014-09-28 03:23:28
