
猴B病毒囊膜蛋白gC基因密码子优化及其在大肠埃希菌中的表达*
2011-03-07 06:13王代平叶华虎白杰英张文韬
动物医学进展 2011年5期
王代平,叶华虎,曾 林,白杰英,董 罡,张文韬,聂 奎
(1.西南大学动物科技学院,重庆400715;2.军事医学科学院实验动物中心,北京100071)
猴B病毒(Monkey B virus),为人兽共患病病原[1]。在自然宿主猕猴体内多呈潜伏感染,临床常见口、生殖器疱疹或呈无症状感染[2],与人感染单纯疱疹病毒(HSV)1型和2型类似。人感染猴B病毒后多表现为脑炎、脑脊髓炎,严重的呼吸麻痹甚至死亡,病死率超过70%,幸存者会留下神经后遗症[3]。
猴B病毒属于生物安全(BSL)4级病原,在自然宿主猕猴中的感染率高(血清学阳性率为10%~60%),并且被感染的猕猴往往终身带毒[4]。临床上多采用与猴B病毒同源的人单纯疱疹病毒(HSV)、猴因子8(SA8)等病毒蛋白作为诊断抗原进行检测,易出现假阳性和假阴性[5]。糖蛋白gC是猴B病毒高度保守的蛋白之一,研究证实gC蛋白ELISA诊断的敏感性97.3%,特异性100%,在猴B病毒4种主要被膜蛋白gB、gC、gD和mgG中,仅次于gB。而gB和gD可以与HSV-1、HSV-2的阳性血清反应。提示gC蛋白可作为猴B病毒诊断的候选抗原[6]。因此,本研究旨在利用密码子优化软件对gC蛋白编码基因序列重新优化设计,替换了大肠埃希菌稀有密码子,采用重叠PCR(overlapping PCR)方法合成目的基因,并高效表达重组gC蛋白,为进一步建立检测猴B病毒抗体间接ELISA方法及研制猴B病毒核酸疫苗奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌种与质粒 原核表达载体pET-28b(+)质粒为美国Novagen公司产品。宿主菌DH5a和Ecoli BL21感受态细胞由西南大学动物科技学院传染病与寄生虫实验室保存。
1.1.2 工具酶和试剂 限制性内切酶EcoRⅠ,HindⅢ,NdeⅠ购自宝生物工程(大连)有限公司。T4连接酶,质粒小提试剂盒购自天根生化科技有限公司。DNA凝胶回收试剂盒购自Qiagen公司。兔抗猴IgG-HRP二抗购自于Sigma公司。猴B病毒标准阳性血清和阴性血清由本实验室保存。
1.2 方法
1.2.1 猴B病毒gC蛋白胞外区编码序列的优化及合成 根据GenBank公布的猴B病毒E2490株gC蛋白的碱基序列,运用密码子优化软件,根据密码子简并性和偏向性原理,在不改变氨基酸序列的前提下,替换大肠埃希菌稀有密码子为高频密码子,在合成片段两端添加NdeⅠ和HindⅢ酶切位点,通过全基因合成,并连接pGM-T载体,测序鉴定。
1.2.2 重组表达质粒pET28b(+)-gC的构建 对pGMT-gC进行双酶切(NdeⅠ和HindⅢ),回收目的片段,插入pET-28b(+)载体,形成重组质粒pET28b(+)-gC,转化DE3大肠埃希菌,经抗性筛选、PCR和酶切等方法鉴定阳性菌落。
1.2.3 重组gC蛋白的诱导表达、纯化及可溶性分析挑选鉴定后的阳性菌落,在含有卡那霉素的LB培养基中培养,至OD600值达0.6左右时,加入IPTG(1 mmol/L)诱导进行表达。SDS-PAGE分析目的蛋白的表达以及表达形式鉴定(分泌型或包涵体)。参照GE Healthcare公司HisPrep FF16/10说明书,进行重组蛋白的纯化。
1.2.4 重组蛋白的Western blot鉴定 纯化后的重组蛋白经SDS-PAGE电泳,以BIO-RAD半干转移系统转膜(PVDF)。用含50 g/L脱脂奶粉的PBS 4℃封闭过夜。PBST洗PVDF膜,分别以50倍稀释的B病毒标准阳性血清37℃孵育1 h;PBST洗膜3×5 min;再以1∶2 000稀释的兔抗猴IgG-HRP孵育,37℃孵育1 h;PBST洗膜3×5min;最后用DAB进行显色。
1.2.5 特异性鉴定 以重组蛋白为抗原,采用方阵实验确定抗原包被浓度及血清工作浓度,辣根过氧化物酶标记兔抗猴IgG以1∶25 000稀释,OPD显色。(结果判定:阳性血清OD490nm值0.5以上,阴性血清OD490nm值0.1以下,P/N值>2.1)。采用ELISA检测方法对HSV-I、HSV-II、BV阳性血清和阴性血清进行特异性检测,同时设阴性和阳性对照。
2 结果
2.1 猴B病毒C蛋白稀有密码子分析结果
猴B病毒gC基因编码区稀有密码子的分布情况见图1,稀有密码子的出现频率高达17%,数据统计见表1。
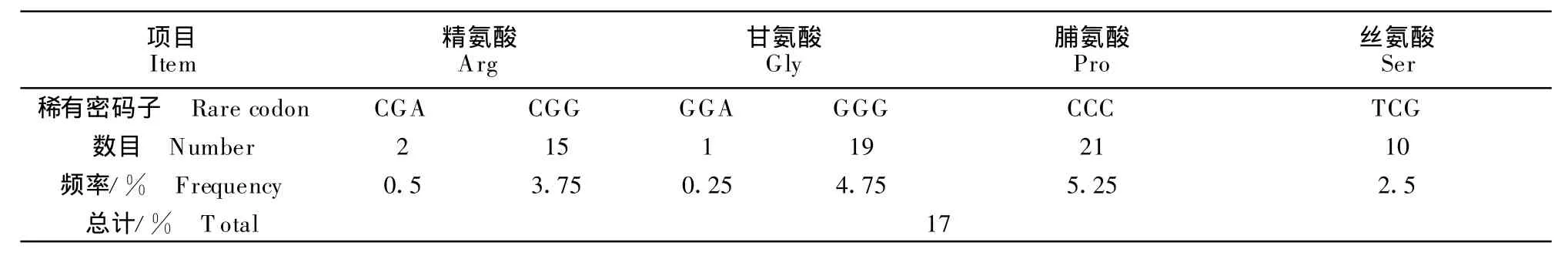
表1 猴B病毒gC基因稀有密码子分析Table 1 Rare codon analysis of monkey B virus-gC gene
2.2 重组表达质粒pET-28b(+)-gC的构建
通过化学合成法获得gC蛋白胞外区序列,片段大小为1 200 bp,与预期大小相符。测序结果表明其序列与优化过的序列完全一致。用NdeⅠ和HindⅢ双酶切pET28b(+)-gC,分别得到约为5 300 bp和1 200 bp的片段,酶切鉴定与预期相符(图1)。
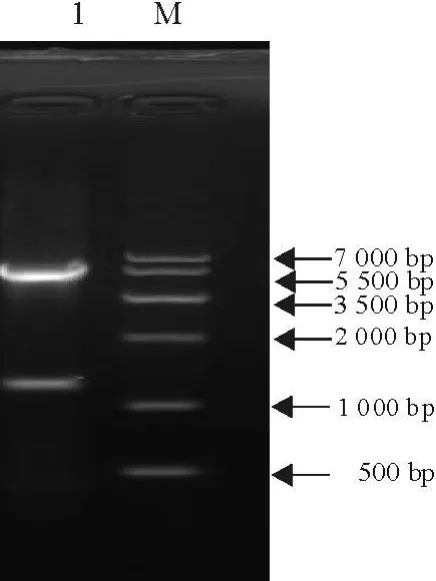
图1 重组质粒pET-28b(+)-gC的双酶切鉴定Fig.1 Enzyme digetion analyses of the recombinant plasmid pET-28b(+)-gC
2.3 重组质粒pET-28b(+)-gC的诱导表达
重组菌诱导后,在约50 ku处出现蛋白条带,与预期重组蛋白的分子质量一致(图2A)。对裂解诱导表达菌株,进一步分析表明,重组蛋白存在于裂解菌沉淀,以包涵体形式存在。凝胶图像分析显示,表达的蛋白约占菌体总蛋白的30%左右。采用Ni-NTA亲和层析柱纯化目的蛋白,SDS-PAGE检测结果表明纯化蛋白为单一条带(图2B),表明纯化效果较佳。
2.4 重组蛋白Western blot鉴定
以猴B病毒标准阳性血清对纯化后的重组蛋白进行Western blot检测,结果显示在大小约50 ku处出现一清晰的特异性反应条带(图3),表明重组蛋白可被猴B病毒阳性血清识别。
2.5 特异性鉴定
将纯化的gC重组蛋白作为包被抗原,经方阵滴定确定抗原最佳包被量为150 ng/孔,血清稀释度为1∶10。以建立的ELISA方法检测与B病毒同属的单纯疱疹病毒的阳性血清,其P/N均小于2.1(表2),表明重组蛋白具有良好的特异性。

图2 SDS-PAGE检测重组蛋白表达Fig.2 SDS-PAGE analysis of recombinant protein
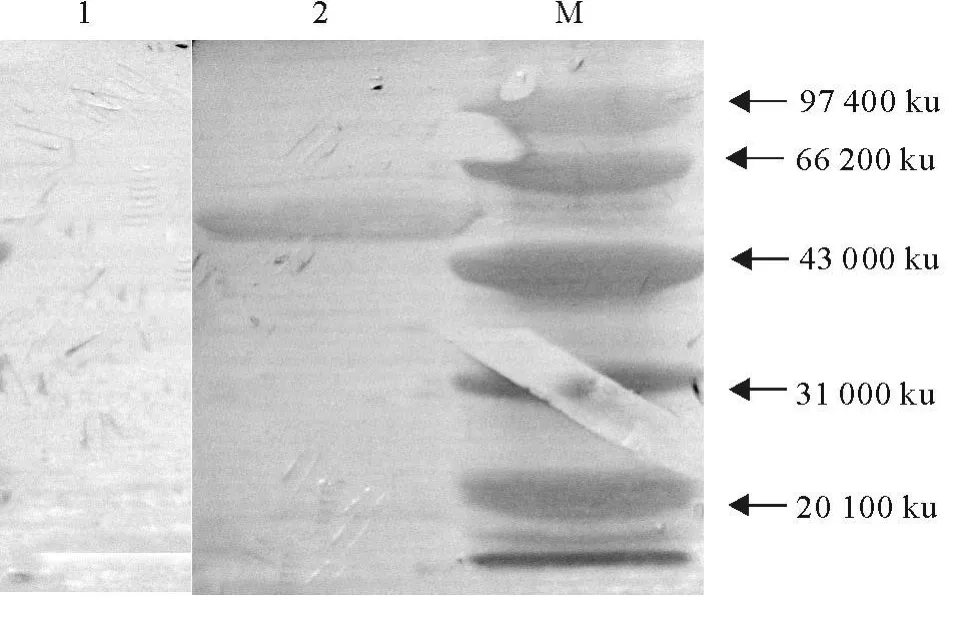
图3 重组蛋白的Western blot分析Fig.3 Western blot analysis of recombinant protein

表2 ELISA检测的特异性试验Table 2 The specificity assay of gC protein by ELISA
3 讨论
本试验最初采用PCR技术以猴B病毒基因组DNA为模板成功扩增出编码gC蛋白胞外区的基因,克隆到表达载体pET-28b(+),鉴定和筛选出含gC胞外区基因的重组质粒,转化表达菌株E.coli BL21(DE3),优化表达条件,未见蛋白表达。之后,我们对糖蛋白gC的胞外区基因片段进行分析,发现含有较多稀有密码子ccc、cgg、gga等,有研究证实,这些稀有密码子在大肠埃希菌中缺乏相应的tRNA,大量或连续出现可能会使蛋白表达量降低甚至不表达[7-8]。我们根据原核细胞对密码子的偏爱性,化学合成糖蛋白gC的胞外区基因序列[9],将其克隆入载体pET-28b(+),转化DE3菌株,优化诱导表达条件获得gC蛋白表达[10]。免疫印迹试验证实,猴B病毒阳性血清和抗His单克隆抗体,能特异性的识别重组蛋白。结果表明,原核表达的猴B病毒gC蛋白具有良好的抗原性。
目前,常用与猴B病毒同源的人单纯疱疹病毒(HSV)作为诊断抗原,这种检测方法,尽管不存在严重的安全隐患,但极大的影响检测的准确性[11]。近几年,国内已有利用基因工程技术构建猴B病毒重组囊膜蛋白,并对其抗原性加以研究的报道,肖镜等[12]表达了猴B病毒gD蛋白片段,并证实重组蛋白具有良好的抗原性。本试验构建的gC重组蛋白,以Western blot验证重组蛋白的抗原性,并初步建立了检测猴B病毒抗体的ELISA方法[13],为有效防控猴B病毒感染提供可行的诊断方法,也为进一步为净化猴群奠定了基础。
[1] Ritchey J W,Payton M E,Eberle R.Clinicopathological characterization of monkey B virus(Cercopithecine herpesvirus 1)infection in mice[J].Comparative Pathology,2005,132(2-3):202-217.
[2] Tregle R W,Loe C L,Earhart R H,et al.Cercopithecine herpesvirus 1 risk in a child bitten by a bonnet macaque monkey[J].J Emerg med,2010,26[Epub ahead of print]
[3] Besecker M I,Harden H E,Li G,et al.Discovery of herpes B virus-encoded microRNAs[J].Virology,2009,83(7):3413-3416.
[4] Huff J L,Eberle R,Capitanio J,et al.Differential detection of B virus and rhesus cytomegalovirus in rhesus macaques[J].Gen Virol,2003,84:83-92.
[5] 韦 毅,符明泰,石寒,等.三种不同抗原对猴B病毒抗体检测结果的比较研究[J].实验动物科学与管理,2001,18(2):1-3.
[6] Perelygina L,Patrusheeva I,Hombaiah S,et al.Production of herpes B virus recombinant glycoproteins and evaluation of their diagnostic potential[J].Clin Microbiol,2005,2(43):620-628.
[7] 杨 丽,郭万柱,殷华平,等.伪狂犬病病毒基因组密码子用法特点分析[J].中国预防兽医学报,2007,29(2):103-106.
[8] Vernet E,Kotzsch A,Voldborg B,et al.Screening of genetic parameters for soluble protein expression in Escherichia coli[J].Protein Expression and Purification,2010,74(2):139-304.
[9] Gvritishvili A G,Leung K W,Tombran Tink J.Codon preference optimization increases heterologous PEDF expression[J].Plot One,2010,5(11):e150-156.
[10] 宫晓炜,周继章.布鲁菌外膜蛋白OMP10表达及其抗原性研究[J].动物医学进展,2009,30(12):25-29.
[11] 王 琼,吴志蕾,艾 军,等.猴B病毒gB基因杆状病毒表达载体的构建[J].云南畜牧兽医,2010(1):4-5
[12] 肖 镜,付 瑞,贺争鸣,等.猴B病毒BVgD-多肽ELISA检测方法的建立[J].实验动物科学,2008,25(2):20-23.
[13] 李 春,宋云峰,金梅林,等.猪流感病毒HA基因的原核表达及其ELISA检测方法的建立[J].动物医学进展,2007,28(7):35-39.
猜你喜欢
生物学通报(2020年11期)2020-10-22
英美文学研究论丛(2018年2期)2018-08-27
中成药(2018年7期)2018-08-04
食品科学(2018年10期)2018-05-23
老年医学与保健(2017年6期)2017-02-06
中国继续医学教育(2015年1期)2016-01-06
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
当代畜禽养殖业(2014年9期)2014-02-27
