
云南猕猴mtDNA控制区全序列测定
2011-02-03 07:38:20禹文海杨凤梅鲁帅尧王俊斌陈丽雄和占龙
中国比较医学杂志 2011年12期
禹文海,黄 芬,杨凤梅,鲁帅尧,赵 远,沈 冬,王俊斌,陈丽雄,和占龙
(1.中国医学科学院北京协和医学院医学生物学研究所,云南昆明 650118; 2.昆明理工大学生命科学与技术学院,云南昆明 650224)
线粒体DNA(m tDNA)是细胞核外遗传物质,呈共价闭合的环状双链结构,能够独立地进行复制和转录,其具有母系方式遗传、碱基变异大、进化速度快等特点。其中,线粒体DNA控制区的进化速度是线粒体DNA其它区域的3~5倍,在生物遗传进化和种群遗传变异程度的研究中应用广泛[1-5]。猕猴又称恒河猴、广西猴、黄猴,为灵长目猴科猕猴属的一个种。云南位于中国的西南方,猕猴资源十分丰富,在北部、西北部、中部、南部和东北部等均有分布[6]。本研究以云南猕猴作为研究对象,测定线粒体控制区全序列,为猕猴的进化分析及开展猕猴遗传资源保护和合理开发利用提供理论依据。
1 材料和方法
1.1 样品采集和DNA提取
实验采集猕猴全血共7份(镇康县1份、景东县1份、镇沅县1份、保山市1份、邓川县1份、牟定县1份、宁蒗县1份),来源于中国医学科学院医学生物学研究所全国医学灵长类研究中心实验动物部,实验动物生产许可证:(滇)SCXK2010—0006。静脉抽取全血2 m L,EDTA抗凝。采用Axygen DNA提取试剂盒提取基因组DNA,详细操作步骤见说明书。
1.2 PCR扩增与目的片段测定
参考相关文献选取引物对[7,8],上游引物F(5'-CCTTACCTGAATTGGAAGCGAACC-3')和下游引物R(5'-GGCCAGGACCAAGCCTATTT-3'),由生工生物工程(上海)有限公司合成。在PCR仪(Bio-Rad C1000)中进行PCR扩增。反应体积为25μL,引物F和R各(20μmol/L)0.5μL,模板DNA 2μL,2× PCR Master M ix 12.5μL,双蒸H2O 9.5μL。反应条件为95℃6 min,然后94℃1 m in,66℃45 s,72℃1 min,40个循环,最后72℃延伸10 min。用1.5%的琼脂糖凝胶电泳检测PCR产物,参照DNA marker DL2000出现1 450 bp左右条带的PCR产物进行纯化测序,测序由上海杰李生物技术有限公司完成。
1.3 序列分析及系统进化树构建
每个个体测得的正反链序列用DNAStar软件进行分析拼接,用Clustalx1.8软件进行序列比对并辅以人工校对,用DNAStar软件对序列间同源性进行分析,用MEGA4.0软件分析序列的碱基组成、转换和颠换。以来源于美国威斯康星州国家灵长类动物研究中心(WRPRC)的猕猴(GenBank No.AY612638)为外群,分别用邻接法(neighbor-joining,NJ)和最小进化法(minimum-evolution,ME)构建分子系统进化树。
2 结果
2.1 序列测定与鉴定
经PCR扩增后得到产物大小约为1 450 bp,去除线粒体DNA控制区两侧的序列后,测得云南猕猴线粒体控制区全序列的长度为1 084~1 089 bp,利用BLAST和GenBank中的猕猴线粒体序列进行比较,同源性达到91.5%以上,说明该序列为猕猴m tDNA控制区。本研究测得的云南猕猴线粒体DNA控制区全序列已提交GenBank数据库,GenBank No.为JF746819(保山)、JF746820(邓川)、JF746824(景东)、牟定(JF834919)、JF746839(宁蒗)、JF746823(镇康)和JF746817(镇沅)。
2.2 序列碱基组成、遗传变异及同源性分析
线粒体控制区核苷酸序列碱基组成见表1,不同地区猕猴线粒体DNA控制区G+C的含量(42.9%~43.8%)低于A+T的含量(56.2%~57.1%),A碱基含量为29.6%~30.2%,C碱基含量为30.4%~31.4%,T碱基含量为26.5%~27.4%,G碱基含量为12.2%~12.5%。序列遗传变异包括转换、颠换、插入和缺失4种形式,经MEGA4.0软件分析可知序列中的转换明显比颠换多,其中T-C转换多于A-G,A-C和A-T颠换多于CG和T-G,转换/颠换比值平均为26.1。
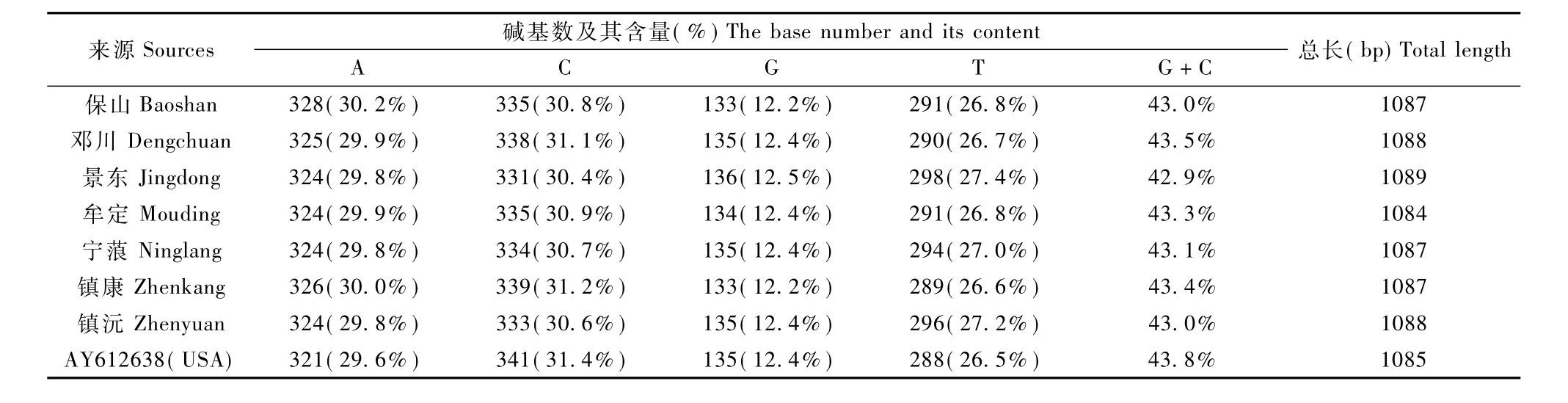
表1 不同地区猕猴线粒体DNA控制区核苷酸碱基组成比较Tab.1 Comparison of nucleotides composition in the m tDNA control region of Macaca mulatta from different areas
不同地区猕猴线粒体控制区全序列间差异及同源性见图1。
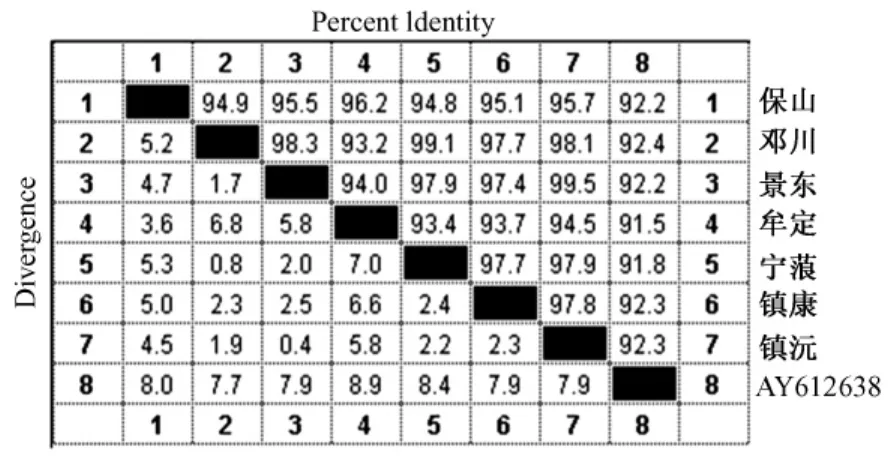
图1 不同地区猕猴线粒体DNA控制区全序列间同源性分析Fig.1 Homology of the m tDNA control region ofMacaca mulatta from different areas
云南不同地区猕猴的同源性为93.2%~99.5%,具有较高的同源性,同源性最高的是镇沅和景东猕猴,其次是邓川和宁蒗猕猴,同源性最低的是邓川和牟定猕猴。与GenBank中参考序列(AY612638)同源性为91.5%~92.4%之间,其中,与邓川猕猴同源性最高(92.4%),与牟定猕猴同源性最低(91.5%)。
2.3 构建系统进化树
以美国威斯康星州国家灵长类动物研究中心的猕猴为外群构建NJ树(图2-A)和ME树(图2-B),各分支上的数值为1000次自举(Bootstrap)分析得到的支持率。从图3中可以看出NJ树和ME树具有相同的拓扑结构,两者均表明所分析的猕猴分为两大分支,来源于美国的猕猴单独为一支,而云南猕猴聚为一支。云南猕猴分为两个平行的姐妹分支,保山和牟定猕猴为一支;景东、镇沅、邓川、宁蒗和镇康猕猴聚为另一支。其中,景东和镇沅以及邓川和宁蒗猕猴遗传关系最近。
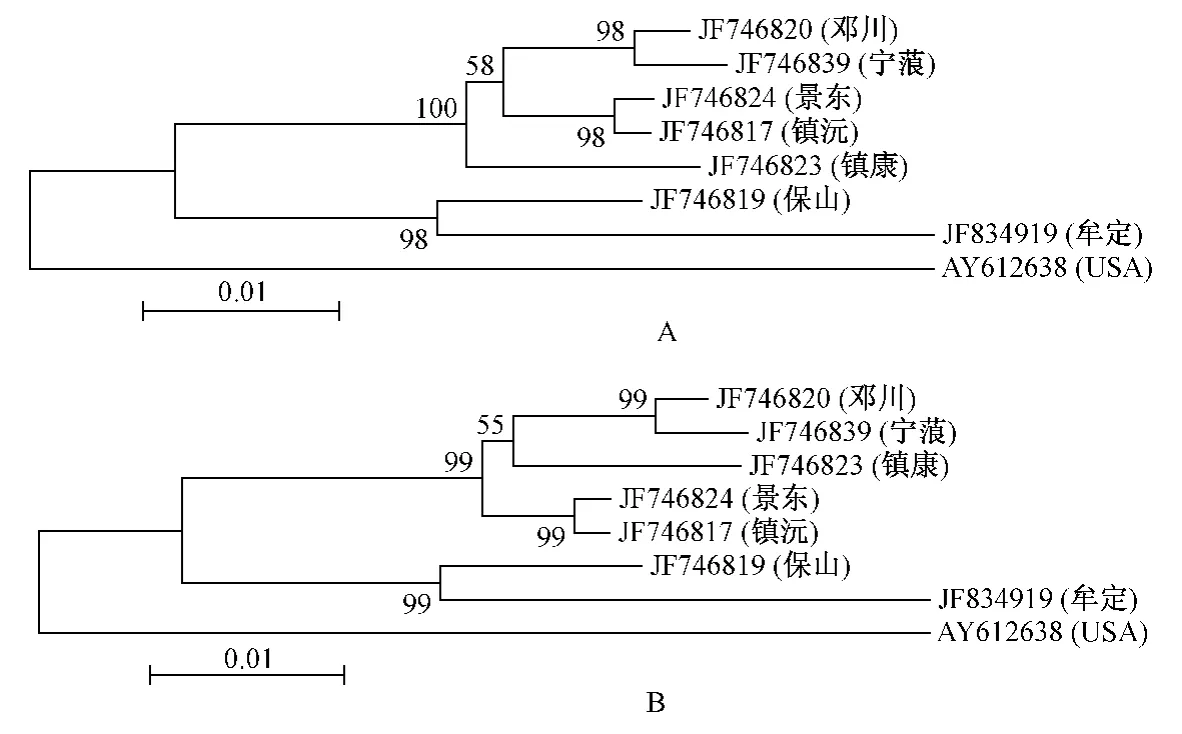
图2 基于线粒体DNA控制区构建的NJ(A)和ME(B)系统树Fig.2 Phylogenetic trees constructed using NJ(A)and ME(B)methodsbased on the comp lete sequences ofm tDNA control region
3 讨论
云南猕猴线粒体DNA控制区全长为1 084~1 089 bp,A、T、G和C四种碱基平均含量分别为29.9%、26.9%、12.3%和30.9%,序列表现出了很强的反G偏倚(即G的含量明显低于其它3种碱基的含量),这与脊椎动物线粒体DNA的G碱基极低现象一致[9-12]。A+T碱基平均含量(56.8%)高于G+C碱基平均含量(43.2%),与其它哺乳动物A、T含量高和G、C含量低的特点相似。序列变异包括了转换、颠换、插入和缺失四种形式,其中转换最为常见,转换与颠换的比值为26.1,明显大于转换与颠换比的临界值2.0[13],表现出较高的转换偏爱性,与其他哺乳动物线粒体DNA序列的研究结果是一致的[14,15],同时也说明云南猕猴线粒体DNA控制区序列突变未达到饱和状态。
我们采用邻接法(NJ)和最小进化法(ME)构建系统进化树,以来源于美国威斯康星州国家灵长类动物研究中心的猕猴为外群,分析不同地区猕猴的遗传进化关系。系统进化树显示云南猕猴聚为一支,而来源于美国的猕猴单独聚为一支。就云南猕猴而言存在两个姐妹分支,来自不同地区的猕猴大多按照其来源地分别相对聚在一起,表现出与地理位置一定的对应关系。目前未见报道国内其它地区猕猴线粒体DNA控制区全序列的测定和提交GenBank数据库,少数研究报道只是测定了其中的部分序列,因此本实验中没有与国内其它地区猕猴群体进行比较研究。
[1]Wenink PW,Baker AJ,Tilanus MG.Mitochondrial controlregion sequences in two shorebird species,the turnstone and the dunlin,and their utility in population genetic studies[J].Mol Biol Evol,1994,11(1):2-31.
[2]潘登,符云新,张亚平.基于线粒体控制区的滇金丝猴群体遗传学研究[J].遗传,2006,28(4):432-436.
[3]柳杨,李进华,赵健元.黄山短尾猴m tDNA控制区序列变异及种群的遗传多样性[J].动物学报,2006,52(4):724 -730.
[4]何丽,张于光,李迪强,等.川金丝猴m tDNA D-loop序列遗传多样性分析[J].动物学杂志,2010,45(1):70-76.
[5]徐怀亮,李地艳,程安春,等.四川地区猕猴线粒体DNA控制区遗传多样性及其种群遗传结构[J].兽类学报,2010,30(3):247-255.
[6]蒋学龙,王应祥,马世来.中国猕猴的分类及分布[J].动物学研究,1991,12(3):241-247.
[7]董高华,徐怀亮,帅素容.藏酋猴线粒体基因组序列测定与进化分析[D].硕士学位论文,2008,四川农业大学.
[8]Blancher A,Bonhomme M,Crouau-Roy B,et al.Mitochondrial DNA sequence phylogeny of 4 populations of the widely distributed cynomolgus macaque (Macaca fascicularis fascicularis)[J].JHered,2008,99(3):254-264.
[9]Sueoka N.Two aspects of DNA base composition:G + C cxnamecontent and translation-coupled deviation from intra-strand rule of A=T and G=C[J].JMol Evol,1999,49(1):49-62.
[10]周继亮,张亚平,黄美华,等.腹亚科蛇线粒体细胞色素b基因序列分析及其系统发育[J].动物学报,2001,47(4):361-366.
[11]杜民,齐兴柱,尹绍武,等.基于Cyt b基因序列研究6种裸胸鳝属鱼类的进化关系[J].中国水产科学,2009,16(1):23-30.
[12]Ding S,Zhuang X,Guo F,et al.Molecular phylogenetic relationships of China Seas groupers based on cytochrome b gene fragment sequences[J].Sci China C Life Sci,2006,49(3):235-242.
[13]Knight A,Mindell DP.Substitions bias,weighting of DNA sequence evolution,and the phylogenetic positions of fea's viper[J].Syst Biol,1993,42(1):18-31.
[14]Gu ZL,Zhao XB,Li N,et al.Complete sequence of the yak (Bos grunniens)mitochondrial genome and its evolutionary relationship with other ruminants[J].Mol Phylogenet Evol,2007,42(1):248-255.
猜你喜欢
小猕猴学习画刊(2022年10期)2022-01-01 04:48:21
天津市教科院学报(2021年5期)2021-11-10 07:32:40
生物学通报(2021年9期)2021-07-01 03:24:44
海洋通报(2020年5期)2021-01-14 09:27:06
童话世界(2020年32期)2020-12-25 02:59:14
小猕猴智力画刊(2019年4期)2019-05-08 21:56:28
小猕猴智力画刊(2019年3期)2019-04-19 00:02:10
中国公路(2017年18期)2018-01-23 03:00:42
水利科技与经济(2017年6期)2017-04-28 08:30:16
汽车纵横(2017年3期)2017-03-18 23:19:33
