
成骨细胞中Runx2对机械离心力刺激的响应
2010-10-13 08:15:30关键程宗生王健平李德超邓慧鑫
华西口腔医学杂志 2010年1期
关键 程宗生 王健平 李德超 邓慧鑫
(佳木斯大学口腔医院 口腔颌面外科,黑龙江 佳木斯 154004)
从Codivilla介绍牵张成骨术(distraction osteogenesis,DO)至今已有一百多年的历史,但目前对牵引产生的机械力如何转化为生物学信号的细胞和分子机制尚不清楚。骨形态发生蛋白(bone morphogenetic protein,BMP)参与DO中新骨的生成并发挥重要作用[1-3],其可能参与DO力学信号向生物信号转化过程[4-5]。BMP信号转导通路在成骨细胞对机械力刺激响应中的作用还不清楚,对该问题的研究将为DO外源性应用BMP提供理论基础。本研究旨在通过生理状态和阻断BMP信号转导通路时,在mRNA层面对比研究成骨细胞株MC3T3-E1中Runx2对机械离心力刺激的响应,从侧面探讨BMP信号转导通路在成骨细胞对机械力刺激响应中的作用和意义。
1 材料和方法
1.1 实验材料
小鼠胚成骨细胞株MC3T3-E1(中国医学科学院基础所细胞中心),DMEM(GIBCO公司,美国),胎牛血清(杭州四季青生物工程材料有限公司),重组小鼠Noggin蛋白(Perpro Tech公司,美国),Trizol(上海生工公司),cDNA逆转录试剂盒、Power SYBRRGreen PCR试剂盒、ABI7300 PCR仪(ABI公司,美国),50 bp DNA Marker(TaKaRa公司,日本),细胞培养瓶/板(Coring公司,美国)。
1.2 MC3T3-E1细胞的培养
37℃、5%CO2、含10%胎牛血清(fetal bovine serum,FBS)的DMEM培养基中培养MC3T3-E1细胞,隔天换液。当细胞铺满瓶底80%以上时,37℃下0.25%胰蛋白酶1.5 mL消化3~5 min,见瓶底出现针孔样透明时,加入等量含10%FBS的DMEM培养基终止消化,吹打细胞制成细胞悬液,按1∶3传代,第2天换液。
1.3 细胞加力
自制六孔细胞培养板离心支架,使之能将匹配的六孔板一起固定于LXJ-Ⅱ型离心沉淀机中离心,离心加力系统的剖面见图1。根据公式:离心力(RCF)=1.118×10-5×R×V2计算离心力的大小,单位为重力加速度×g。离心半径R约19 cm,转速V约1 100 r·min-1,离心力约271×g。
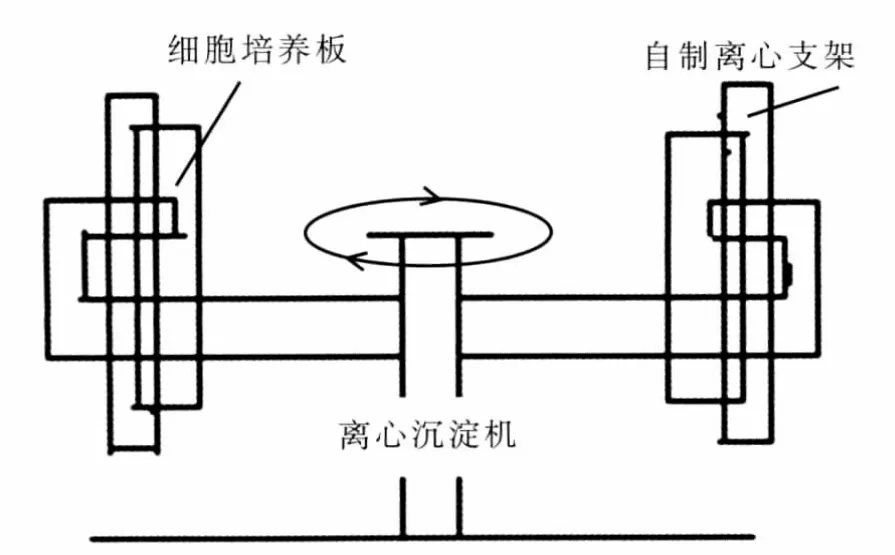
图1 离心加力系统的剖面图Fig 1 Centrifugal profiles based augmentation systems
1.4 加力分组
将待测细胞随机分为A、B、C、D组。A组为空白对照组,细胞未接受Noggin预处理和机械离心力加载;B组为细胞接受271×g离心力加载5 min;C组为细胞接受100 ng·mL-1Noggin预处理24 h;D组为细胞接受100 ng·mL-1Noggin预处理24 h和271×g离心力加载5 min,每组6孔。4组细胞同时种板,培养48 h后以无血清的DMEM继续培养24 h,同步细胞周期。加力前24 h更换为含10%FBS的DMEM培养基,将六孔板无菌密封。30min后4组分别收获细胞。
1.5 Runx2表达的检测
按Trizol试剂说明提取细胞总RNA,并逆转录成cDNA。采用Power SYBR®Green染料法进行PCR,引物由Invitrogen公司合成,目的基因Runx2(Gene-Bank accession number NM_004348)上游引物序列为:5′-TCCAACCCACGAATGCACTAC-3′,下游引物序列为:5′-GTAGTGAGTGGTGGCGGACT-3′,扩增产物片段为92 bp;内参对照β-actin(GeneBank accession number NM_007393)上游引物序列为:5′-CATCCGTAAAGACCTCTATGCCAAC-3′,下游引物序列为:5′-ATGGAGCCACCGATCCACA-3′,扩增产物片段为171 bp。PCR反应条件为:95℃ 10 min,95℃ 15 s,62℃ 1 min,共40个循环。PCR产物同期进行琼脂糖凝胶电泳分析。
1.6 统计学分析
2 结果
2.1 Runx2对离心力刺激的响应
MC3T3-E1细胞中Runx2 mRNA的检测结果见图2。由图2可见,B组Runx2 mRNA表达较A组增加(P<0.05),离心力可使Runx2 mRNA表达增加。D组经Noggin预处理后比B组对离心力刺激时Runx2 mRNA表达降低(P<0.05),Noggin抑制Runx2 mRNA的表达。C组Runx2 mRNA表达无明显变化,A、C、D组间差异无统计学意义(P=0.692)。

图2 成骨细胞Runx2 mRNA的表达Fig 2 The expression of Runx2 mRNA in osteoblastic MC3T3-E1 cells
2.2 实时荧光定量PCR产物电泳分析
凝胶电泳结果显示PCR产物杂带少,RT-PCR产物纯度较高(图3)。
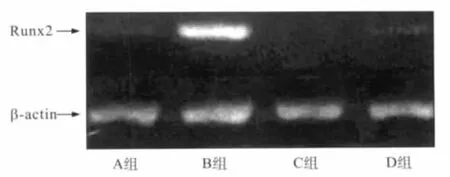
图3 Runx2表达产物凝胶电泳图Fig 3 The electrophoresis results of expression product of Runx2
3 讨论
成骨细胞是骨组织内的力学敏感性细胞,同时它在骨改建过程中又处于一个中心调控的地位,是研究骨形成机制的基本单位[6-7]。MC3T3-E1细胞表型稳定,使其在对成骨细胞的各项研究中得到了广泛地应用。目前对离体细胞有多种加力方式:低渗性膨胀、离心、单/双向拉伸和流体剪切等,这些不同的离体应变使细胞产生不同的生理响应过程。本实验的离心加力方式与Fitzgerald等[8]所提出的类似,离心力使贴壁生长的细胞受到一个顶—底轴向的压力,避免了细胞悬浮离心时细胞之间以及细胞与基质之间附着状态的丧失。Fitzgerald等[8]发现对MC3T3-E1细胞加载3×g的离心力相当于轻微力量,而287×g的离心力则相当于剧烈刺激的力量。前期实验表明对MC3T3-E1细胞加载271×g的离心力5 min能有效刺激成骨细胞,并且离心力刺激强度在成骨细胞生理范围之内。离心力在一定程度上反映成骨细胞受到机械力学刺激作用,但是无法完全模拟体内DO成骨细胞的实际受力情况。
成骨细胞对力学信号的转导通过胞膜上Ca2+通道、胞外基质-整合素-细胞骨架结构系统[9]、细胞因子等多种途径入核作用于靶基因Runx2并调控相应基因表达构成复杂的力学信息传递网络。本研究结果显示:与A组相比,B组Runx2 mRNA表达增加,可能为成骨细胞对机械离心力刺激响应的结果,验证了Runx2作为力学信号靶分子参与了成骨细胞响应机械离心力刺激的生理过程。Ziros等[10]证实Runx2/Cbfa1蛋白及基因都是力学刺激的关键目标,认为Runx2/Cbfa1基因表达的增强是力学刺激在成骨细胞内作用的“终点”。Li等[11]对成骨样细胞施加离心力10 min后检测到Runx2/Cbfa1 mRNA表达水平有短暂地增加改变,认为成骨细胞通过多种复杂途径响应机械力学刺激。
采用Abe等[12]的方法阻断BMP信号转导,使用100 ng·mL-1Noggin预处理24 h,其Runx2 mRNA表达水平可反映Noggin阻断BMP信号转导通路的效果,结果显示,C组与A组和D组间差异无统计学意义,认为D组达到了阻断BMP信号转导通路的要求。D组Runx2 mRNA表达水平略高于A组,可能是除BMP信号转导通路外,其他信号通路对机械离心力刺激响应的效果。
D组Runx2 mRNA表达水平较B组显著降低。仅阻断BMP信号转导通路,从现有理论来讲,其他多种力学信号传递途径应不受影响,但是BMP信号转导通路阻断后,力学靶点Runx2基因在相同的力学刺激强度下未能完成与B组同等的响应程度,二者对机械离心力刺激响应程度的差异具有统计学意义(P<0.05)。提示BMP信号转导通路参与了成骨细胞对机械离心力刺激的生理响应过程,而且可能在成骨细胞对力学信号引起的信息传递级联反应中扮演了重要角色。力学刺激可能通过BMP/Smad途径[13-15]或BMP/p38MAPK途径[16-17]等入核作用于Runx2等影响成骨特异性基因表达,从而刺激骨形成以响应力学刺激,但其具体机制尚不清楚。BMP信号转导通路在成骨细胞对机械力刺激响应中发挥了重要作用。BMP信号转导通路可能介导力学信息传递网络的其他途径,并在力学刺激引起的信息传递级联反应中扮演重要角色,但还需要进一步研究验证。
[1]Yonezawa H,Harada K,Ikebe T,et al.Effect of recombinant human bone morphogenetic protein-2(rhBMP-2) on bone consolidation on distraction osteogenesis:A preliminary study in rabbit mandibles[J].J Craniomaxillofac Surg,2006,34(5):270-276.
[2]Zheng LW,Wong MC,Rabie AB,et al.Evaluation of recombinant human bone morphogenetic protein-2 in mandibular distraction osteogenesis in rabbits:Effect of dosage and number of doses on formation of bone[J].Br J Oral Maxillofac Surg,2006,44(6):487-494.
[3]陈安威,魏奉才,王克涛,等.腺病毒介导的人骨形成蛋白2基因转染促进下颌牵张成骨的实验研究[J].中华口腔医学杂志,2007,42(1):15-17.
CHEN An-wei,WEI Feng-cai,WANG Ke-tao,et al.Effect of bone morphogenetic proteins-2 on mandibular distraction osteogenesis in rabbits[J].Chin J Stomatol,2007,42(1):15-17.
[4]Campisi P,Hamdy RC,Lauzier D,et al.Expression of bone morphogenetic proteins during mandibular distraction osteogenesis[J].Plast Reconstr Surg,2003,111(1):201-208.
[5]Khanal A,Yoshioka I,Tominaga K,et al.The BMP signaling and its Smads in mandibular distraction osteogenesis[J].Oral Dis,2008,14(4):347-355.
[6]关键,王健平,车笑非,等.缺氧条件对成骨细胞VEGF和TGF-β表达影响的实验研究[J].中国修复重建外科杂志,2008,22(8):984-988.
GUAN Jian,WANG Jian-ping,CHE Xiao-fei,et al.Experimental study on the effect of hypoxia on the expression of VEGF and TGF-β1 of rat mandibular osteoblasts[J].Chin J Reparat Reconstr Surg,2008,22(8):984-988.
[7]关键,阚娜,刘凤玲,等.大鼠成骨细胞体外培养、特性鉴定及低氧时VEGF表达的变化[J].中国口腔颌面外科杂志,2007,5(6):451-455.
GUAN Jian,KAN Na,LIU Feng-ling,et al.In vitrostudy of expression of VEGF in osteoblast in Wistar rat under hypoxia[J].Chin J Oral Maxillofac Surg,2007,5(6):451-455.
[8]Fitzgerald J,Hughes-Fulford M.Mechanically induced c-fos expression is mediated by cAMP in MC3T3-E1 osteoblasts[J].FASEB J,1999,13(3):553-557.
[9]Carvalho RS,Bumann A,Schaffer JL,et al.Predominant integrin ligands expressed by osteoblasts show preferential regulation in response to both cell adhesion and mechanical perturbation[J].J Cell Biochem,2002,84(3):497-508.
[10]Ziros PG,Gil AP,Georgakopoulos T,et al.The bone-specific transcriptional regulator Cbfa1 is a target of mechanical signals in osteoblastic cells[J].J Biol Chem,2002,277(26):23934-23941.
[11]Li J,Jiang L,Liao G,et al.Centrifugal forces within usuallyused magnitude elicited a transitory and reversible change in proliferation and gene expression of osteoblastic cells UMR-106[J].Mol Biol Rep,2009,36(2):299-305.
[12]Abe E,Yamamoto M,Taguchi Y,et al.Essential requirement of BMPs-2/4 for both osteoblast and osteoclast formation in murine bone marrow cultures from adult mice:Antagonism by noggin[J].J Bone Miner Res,2000,15(4):663-673.
[13]Derynck R,Zhang YE.Smad-dependent and Smad-independent pathways in TGF-beta family signaling[J].Nature,2003,425(6958):577-584.
[14]Miyazono K,Maeda S,Imamura T.BMP receptor signaling:Transcriptional targets,regulation of signals,and signaling crosstalk[J].Cytokine Growth Factor Rev,2005,16(3):251-263.
[15]Wan M,Cao X.BMP signaling in skeletal development[J].Biochem Biophys Res Commun,2005,328(3):651-657.
[16]程宗生,关键.骨形态发生蛋白/Smad信号转导机制及其对离心力刺激的响应[J].国际口腔医学杂志,2009,36(3):335-337.
CHENG Zong-sheng,GUAN Jian.Mechanism of bone morphogenetic protein/Smad signaling pathway and its response to centrifugal force[J].Int J Stomatol,2009,36(3):335-337.
[17]Higuchi C,Myoui A,Hashimoto N,et al.Continuous inhibition of MAPK signaling promotes the early osteoblastic differentiation and mineralization of the extracellular matrix[J].J Bone Miner Res,2002,17(10):1785-1794.
猜你喜欢
宁夏医科大学学报(2022年6期)2022-11-22 08:33:29
云南医药(2021年3期)2021-07-21 05:40:20
临床肝胆病杂志(2017年1期)2017-03-07 03:04:26
安徽医科大学学报(2016年12期)2017-01-15 14:21:53
中国民族医药杂志(2016年6期)2016-05-09 08:52:52
幼儿教育·父母孩子版(2015年2期)2015-05-30 10:48:04
中国医科大学学报(2015年10期)2015-03-01 02:09:58
中国医药导报(2015年24期)2015-02-28 22:07:30
现代检验医学杂志(2014年2期)2014-02-02 02:40:54
中国医学科学院学报(2011年5期)2011-12-01 03:52:51
