
马铃薯病毒研究新进展
2010-09-12 13:36马雪青王永刚周贤婧赵虎彪李昆鹏马建忠
食品工业科技 2010年10期
马雪青,王永刚,周贤婧,赵虎彪,李昆鹏,马建忠
(兰州理工大学生命科学与工程学院,甘肃兰州730050)
马铃薯病毒研究新进展
马雪青,王永刚,周贤婧,赵虎彪,李昆鹏,马建忠*
(兰州理工大学生命科学与工程学院,甘肃兰州730050)
马铃薯病毒作为影响马铃薯产量的主要因素,备受研究者的青睐。国际上对马铃薯病毒结构、检测手段的研究,逐渐从病毒形态学研究转到病毒基因组学及功能组学的研究。伴随着测序技术的不断发展,目前已有14种马铃薯病毒全基因测序完成,对马铃薯病毒种属鉴定、检测技术、基因结构和功能、复制与转录、传播及抗病毒的研究有了最新的进展。本文就上述研究给予总结,对14种马铃薯病毒进行了进化树分析,同时对马铃薯病毒蛋白质组学和代谢组学进行展望。
马铃薯病毒,检测,基因结构与功能,进化
Abstract:In recent years,potato virus as a main factor affecting potato yield are in the favor of researchers.Internationally potato virus have advanced from virus morphology study to genomics and functional genomics research.With the sequencing technology development,14 species of potato virus have had complete genome sequences now.And there are new advances in the potato virus research on species identification,detection,gene structure and function,the mechanism of replication and transcription,transmission and anti-virus and so on.This paper gives a summary of these studies,analyses the homology for 14 kinds of potato virus by phylogenetic tree,and gives an overview of the researches on proteomics and metabolomics applications in potato virus.
Key words:potato virus;detection;gene structure and function;evolutionary
马铃薯作为世界第四大粮食作物,仅次于小麦、水稻、玉米。它主要是通过块茎无性繁殖,其产量在很大程度上取决于马铃薯“种子块茎”的质量,但是由于马铃薯病毒的存在使其产量大大降低,直接造成巨大的经济损失。目前已知的感染马铃薯的病毒有40余种[1],随国家、地域分布而不同。造成世界范围内马铃薯产量下降的马铃薯病毒主要有马铃薯卷叶病毒(potato leaf roll virus,PLRV)、马铃薯Y病毒(potato virus Y,PVY)(包括 NNTN株系)、马铃薯X病毒(potato virus X,PVX)、马铃薯S病毒(potato virus S,PVS)、马铃薯M病毒(potato virus M,PVM)、马铃薯A病毒(potato virus A,PVA)和纺锤块茎类病毒。因此作为世界主要经济粮食作物之一,世界各国都建立自己的马铃薯研究院所,譬如设立在秘鲁利马隶属于国际农业研究组(农研组CGIAR)的16个国际农业研究中心的协商小组之一的国际马铃薯研究中心(CIP)、英国马铃薯理事会、美国马铃薯协会等。马铃薯病毒的研究也成为各国研究的重点,马铃薯病毒的检测技术愈来愈成熟、快捷、灵敏,同时某些马铃薯病毒的分子生物学用途也在引起科研单位的重视,并且先后对病毒进行亚单位乃至全序列进行了分析、克隆、构建与表达等,马铃薯抗病毒植物基因工程也在不断的研究中。
1 马铃薯病毒检测方法比较
自植物病毒发现以来,世界各地科学家们都在绞尽脑汁的思考适合的检测方法。事实上,确实建立了许多适宜的检测方法,能够准确鉴定各种病毒,主要涉及到生物学测定、血清学测定、生理生化测定、电镜技术、分子生物学技术等多种检测手段。病毒的检测方法要求迅速、准确、廉价。尤其是马铃薯作为世界第四大粮食作物,不断遭受病毒的侵害,产量不断下降。因此建立适合于马铃薯病毒的检测方法也迫在眉睫,2006年Khaled Makkouk针对植物病毒检测方法给了精确描述[2]。总体上可以归纳为三大类:生物学方法;免疫学方法;核酸介导的分子生物学方法以及各种方法的综合应用。
1.1 生物学方法
生物学方法是植物病毒研究工作的基本环节,亦是准确诊断鉴定病毒的关键,曾在马铃薯植物病毒鉴定与检测工作中起着重要作用。马铃薯病毒生物学检测常规内容主要包括寄主范围测定、鉴别寄主反应、病毒汁液钝化温度、稀释限点、体外存活期等离体性状测定和传播途径测定等,虽费时、费事,但对马铃薯病毒的生物学特性早期鉴定起到重要的作用。马铃薯病毒的生物学检测策略见图1。2003年吴凌娟[3]利用指示植物,通过接种、培养从形态学角度对马铃薯PVX的寄主范围、症状表现进行了研究,文中充分体现了传统分类鉴定方法的研究策略,虽然繁冗,但对早期病毒的形态学认识起到很重要的作用;2000年吕典秋[4]等通过摩擦接种马铃薯病毒PVX于普通烟上作为繁殖植物,利用PEG沉淀和差速离心技术分离提纯了马铃薯病毒PVX;从一定角度对病毒的传播途径以及分离提纯方法给予介绍。S.M.Paul Khurana[5]亦对马铃薯Y病毒的分类、传播方式给予介绍。从中我们可以了解到PVY为马铃薯Y病毒属数目最多的病毒,病毒基因变异是导致数目巨大的根本原因,而病毒传播又是导致病毒基因重组及变异的主要原因。ICTV(The Universalvirus Datebase of the International Committee on Taxonomy of viruses)收录了源于世界各地关于马铃薯病毒的研究,并且从起源、分类、形态学特征、生理生化性质等方面给予了叙述。自从1939年Kausche第一次应用电子显微镜把烟草花叶病毒粒体照相以后,电镜技术的研究也突然增加起来,针对马铃薯病毒的电镜图片也随之而来,见图2[6],使得对马铃薯病毒形态结构特征有了清楚的认识。马铃薯病毒生物学鉴定方法的应用使得人们从宏观角度认识到病毒的基本特点、生理性质以及传播途径等,在一定角度上有助于人类进行病毒防御,间接提高了马铃薯等粮食作物的产量。
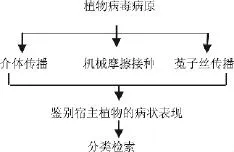
图1 病毒生物学检测策略
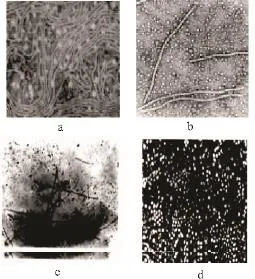
图2 马铃薯病毒的电镜图片
1.2 免疫学方法
免疫学测定方法不仅可以用于病毒分类的鉴定,还可定量地测定病毒在寄主体内的分布,如马铃薯的几种重要病毒的抗血清曾被用来测定马铃薯块茎的带毒率[7]。血清学反应主要是病毒外壳蛋白抗原决定簇的作用,将有血清关系的病毒归在一个属内,但是在同一病毒属内也并不是所有病毒之间都有血清学关系,马铃薯Y病毒属病毒之间血清学关系就比较复杂。免疫学测定的方法有沉淀法、凝结法、免疫扩散法、酶联免疫吸附法、斑点免疫法、免疫电泳和免疫电镜法等。目前主要集中在酶联免疫吸附法ELISA(Enzyme-linked immunosorbent assay)及其衍生方法如TIBA(Tissue blot immunoassay)、Quartz crystal microbalance(QCM)immunosensors等方面[8]。郭兴启等[9]报道了马铃薯Y病毒两分离物的生物学和血清学特性、病毒粒体形态大小以及它们引起的寄主细胞病变特征等方面的差异。周艳玲[10]等论述了利用ELISA等方法对马铃薯Y病毒属的检测。R.P.Singh等论述了利用血清学方法分离鉴定了重组马铃薯PVY病毒[11]。刘卫平等[12]采用快速法检测了马铃薯PVX、PVY病毒;白艳菊等[13]应用快速DAS-ELISA同时检测PVX、PVY、PVS、PVM、PLRV5种马铃薯病毒。常规ELISA中各种反应是在静止状态下进行的,而快速ELISA中各种反应是在振摇状态下进行的。快速法和常规法具有相同的阳性率和灵敏度,但精密度高于常规法,且所用时间远远少于常规法。1991年张国柱[14]用Dot-ELISA对马铃薯主要病毒PVX、PVY、PLRV进行了检测,1993孟清等[15]用这种方法检测了马铃薯PVX、PVY和PVS病毒。相较生物学鉴定方法,免疫学方法灵敏度高、特异性强、操作简单、适用于大量田间样品的检测,是目前生产应用最便捷、使用最为广泛的检测技术。
1.3 核酸介导的分子生物学方法以及各种方法的综合应用
以核酸介导的分子生物学病毒检测方法主要建立在PCR(polymerase chain reaction)反应基础上,包括RT-PCR(Reverse transcription-polymerase chain reaction)技术、核酸杂交(Nucleic acid hybridization)技术、测序技术等。主要步骤有核酸RNA的提取;引物设计与扩增、克隆;测序鉴定;杂交分析。RT-PCR检测技术在马铃薯病毒分子检测技术中应用最广,具有简单、快速、灵敏、特异性强、重复性好等优点,仅用微量染病组织的汁液即可灵敏地检测到病毒的存在。此类技术不仅可以在基因水平上为植物病毒的检测提供更灵敏的手段,而且可与核酸序列分析结合,检测基因序列的分子变异,分析病毒株系间的序列同源性比较亲缘关系,为病毒的鉴定提供可靠依据。
目前该项技术已分别应用于PVY、PVA、PLRV、PVS、PVX等病毒的检测中。2009年Hugh Barker等通过RT-PCR方法克隆了PVY病毒部分基因片段、同时利用测序、分析等手段鉴定了6种PVYO,PVYN和PVYNTN亚种病毒,有助于从基因到结构功能的进一步认识[16]。2006年J.Aramburu等对源于西班牙东北部39种PVY利用生物学、血清学及分子生物学手段进行鉴定,得出此类病毒的基因型多样性、属于不同亚种的结论[17]。2005年Ismail Abdullahi等对寄生在马铃薯体内病毒18S rRNA基因进行克隆、序列分析;建立了能同时检测多种病毒的核酸微矩阵方法[18]。2000年K.-H.Pastrik利用多重PCR方法快速准确鉴定了马铃薯块茎亚种病毒subsp.sepedonicus[19]。1999年L.Glais等根据病毒基因组5′末端序列多态性特点,利用PCR-RFLP(restriction fragment length polymorphism)方法鉴定PVY病毒亚种病毒,得出引起马铃薯块茎坏死病毒PVYNTN属于同一亚种的结论[20]。2002年叶明等[21]以及1998年H-L WEIDEMANN分别应用单酶-RT-PCR、RT-PCR方法对马铃薯纺锤体病毒进行了鉴定[22]。
总之,马铃薯病毒检测技术经历了传统生物学检测技术、免疫学检测技术和分子生物学检测技术三个阶段。传统生物学检测技术准确、直观、易于操作,但耗时长,已不能适应生产需要。免疫学检测技术快速、灵敏,适用于大量样品的检测,但传统免疫学技术中在抗血清的制备上还有很多不足。随着分子生物学技术的不断发展,一些技术应用到马铃薯病毒检测中将是必然趋势。随着RT-PCR技术及相应仪器的普及,将成为应用最广泛、最有前途的病毒检测技术。此外分子生物学技术与免疫学技术的结合推动了两种技术的共同发展,直接通过单链抗体片段的筛选及抗体基因克隆及表达即可完成抗血清的制备,解决了传统免疫学技术中的难以解决的抗原制备问题。能快速准确地鉴定马铃薯病毒分子。
2 马铃薯病毒的分子生物学研究
自从上世纪五十年代末期至今,马铃薯病毒的研究从一般的生物化学及生物物理的分析研究逐渐扩展到分子生物学领域的研究。一方面研究马铃薯病毒的形态学特征、生理生化性质、病毒蛋白和核酸的结构组成;另一方面又研究了核酸及蛋白的复制、转录、翻译和侵染机制。同时,在此基础上又不断的衍生出针对马铃薯抗病毒的研究,包括马铃薯组织培养技术的研究、脱毒技术的研究、抗病毒基因工程马铃薯的培育等。总之集中体现在三方面的研究基因结构及功能分析;生物系统进化分析;病毒传染机制及抗病毒转基因工程研究。
2.1 基因结构及功能分析
马铃薯病毒的研究主要经历了从宏观形态学到微观病毒基因复制、转录、翻译机制以及结构功能研究两个阶段。病毒颗粒主要由核酸及蛋白衣壳两部分构成,少数病毒衣壳部分还包括脂质。人们为了解析病毒结构主要从核酸和蛋白两部分进行研究。一方面利用物理学手段,另一方面主要通过病毒基因的克隆与表达,来解析蛋白的结构与功能。2009年Hui quan Liu等对病毒基因及结构研究策略给予系统介绍[23],见图3。2002年Tuija Kekarainen等通过构建基因文库方法,研究了PVA病毒的基因组序列、结构及各部分功能,指出PVA结构包括5′NTR、HC-Pro、P3、C1、VPg、Nia-Pro、Nib、CP、3′NTR九部分,同时绘制了Genome-Wide Map of Sites Essential图谱[24]。2008年 Amy Kendall等利用光纤衍射、低温电子显微技术、扫描透射电镜进一步阐述了PVX的螺旋对称性结构、核酸及蛋白的排布结构,蛋白亚基结构等[25]。2006年Xuehua Zhong等利用Isostericity Matrix及突变手段研究了马铃薯块茎病毒(PSTV)RNA病毒的三级结构和基序,绘制了PSTV的二级结构及病毒复制过程图谱,阐述了三级结构在病毒复制过程中所起的关键性作用[26]。
此外,分子克隆技术的日益成熟,使得人们不仅对病毒基因的结构与功能有了更深层次的理解,同时对病毒的复制转录机制也有了清晰的认识。2004年Richard H.Guenther等利用RNA基序矩阵、NMR以及计算机软件分析等手段分析了RNA病毒的结构、对亚结构进行了结构与功能分析,指出RNA病毒的复制与转录是通过个别基序与核酸、蛋白相互作用来介导[27]。总之,依赖于RNA聚合酶的RNA病毒的复制、转录机制研究目前普遍认为由病毒自身基因组中的顺式作用原件决定的[28-29]。2007年Victoria S.Korneeva和Craig E.Cameron研究了依赖于RNA聚合酶的病毒结构和功能间的关系,文中指出了病毒复制的忠实性、复制速度以及起始机制是由与核糖体结合部位的核苷酸残基决定的[30]。2009年Helena Plchova,Noemi Cerovska等首次对源于捷克的VIRUBRA1/045(EU717545)、VIRUBRA 1/046(EU717546)和VIRUBRA 1/047(EU313202)PLRV基因组序列进行了分析,同时跟NCBI收录的13种不同宿主的PLVR全基因组序列进行了比对分析,得出最高一致性为98.7%。指出PLRV基因组包括7个ORF和1个RAP,编码8种蛋白,负责病毒复制、侵染、繁殖过程中的蛋白质装配、衔接等作用,其中ORF 3和ORF 4为保守序列、ORF 0和RAP为可变序列[31]。
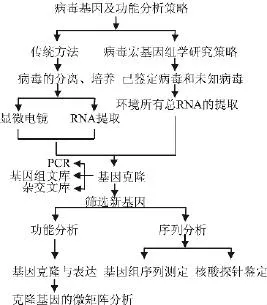
图3 病毒基因及结构分析策略
2.2 生物系统进化分析
植物病毒的生物学分类迄今为止仍然是科学研究领域的一大难题,传统的生物学鉴定方法不能快速准确地进行种属鉴定。而且其进化关系的分析也比较困难,就马铃薯病毒而言,其实很早就存在,但是对它们的认识却经历了较长的时间,而且部分马铃薯病毒分类很难界定,给其病理学研究带来了一定困难。自从分子生物学方法的不断更新、成熟,使得这一难题能够简便、快速、准确的解决。目前许多研究人员已经克隆分析了来源不同的多种马铃薯病毒,在the National Center for Biotechnology Information(NCBI,http://www.ncbi.nlm.nih.gov/)数据库内可以检索到1968种马铃薯病毒基因序列。为了分析其进化关系,可以从许多方面出发,总之要选取保守性强的片段进行比对分析建树。2004年G.Stuart等从NCBI中挑选25种蕃茄丛矮病毒科病毒,分别选取聚合酶、病毒外壳基因和编码外壳运动蛋白基因作为基准,进行比对分析,推测和鉴定了蕃茄丛矮病毒科病毒的发展进化史,同时对基因结构进行了系统分析,得出了准确的编码功能蛋白的基因结构[32]。本文将14种已经得到的完整基因组序列的马铃薯病毒利用CLC DNA Workbench 5.0软件进行进化树分析,得到基于马铃薯病毒全基因序列的Bootstrap Neighbor-Joining进化树,见图4。从图中我们可以看出其种属以及种属间的进化距离等,能清楚地推测其发展进化史。

图4 基于各种马铃薯病毒全基因序列构建的Bootstrap Neighbor-Joining进化树
2.3 马铃薯病毒侵染机制及抗病毒转基因研究
2.3.1 病毒侵染机制的研究 马铃薯病毒和其他侵染性的病原物一样,由于它的寄生性及致病性,破坏人类赖以生存的农作物,马铃薯病毒结构简单,而且有些病毒颗粒在结晶以后,尚能保持其繁殖能力,许多生物学家及生理学家都集中精力来研究其侵染机制,希望借此来破坏其侵染等特性。
目前普遍认为病毒的传播主要是靠蚜虫传播,而其侵染植株主要是靠病毒颗粒的运动性引起的,而蚜虫的传播又与病毒颗粒的结构有关系,2008年Mónica Betancourt等指出PVY病毒的传播除了要靠蚜虫的传播,同时还取决于PVY的结构,其作为结构简单病毒相较结构复杂病毒更加容易被蚜虫所携带,进行传播。而病毒颗粒一旦进入健康植株,其侵染健康植株的特性又是由靠病毒在细胞间的运动所需要的自身编码的运动蛋白来完成[33]。2003年Konstantin I.Ivanov等利用蛋白激酶CK2对Potyvirus属PVA病毒外壳蛋白磷酸化,体外侵染植物,发现磷酸化作用在病毒侵染过程中其重要作用[34]。2002年Natalia E.Yelina等研究发现PVA病毒的长距离运动是靠自身编码的运动蛋白Hcpro协助完成的[35]。2001年O.N.Fedorkin等采用突变技术对PVX中编码外壳蛋白C端18个核苷酸残基进行逐渐缺失,通过克隆突变基因,在烟草中表达分析。分析得出C端核苷酸残基编码蛋白质在病毒细胞间的运动过程中起关键性作用[36]。
马铃薯病毒属病毒至今未确定专一的运动蛋白,有研究者通过病毒编码区基因的突变体研究其在宿主体内的运动,研究表明突变体病毒粒亦可在植株体内运动,猜测运动蛋白的形成与宿主植物自身的结构组成有关系,植株体内的某些蛋白协助病毒颗粒运动。而且前者理论较为流行,因此要解除破坏其侵染特性,主要是靠破坏其运动性来完成。运动蛋白主要分布在外壳上,编码外壳蛋白的基因的研究自然而然又成为研究焦点。世界各地研究人员相继从不同来源的带病植株体内克隆到病毒外壳蛋白基因,以期从根本上了解其性质,解除病毒侵染破坏性。2003年V.Brault等利用点突变的方法将编码枯黄病毒外壳主要蛋白的7个S domain区进行突变,进而研究不同domain编码蛋白对病毒颗粒的沉积、蚜虫介导传播的影响作用,研究表明病毒的侵染运动是靠各类蛋白相互协同作用完成的[37]。
此外也有人推测病毒运动蛋白可能参与胞间运输,2007年Daniel Hofius等采用了酵母双杂交系统方法、定点突变、绿色蛋白融合表达手段研究发现辅助PVY细胞间运动以及长距离运动是由自身基因的编码运动蛋白和宿主细胞内某些蛋白的协同完成的。同时指出运动蛋白核心区的蛋白部分NtCPIPs在病毒的形成、运动过程中对胞间连丝的形成、沉积、侵染等方面起决定性作用[38]。
2.3.2 抗病毒转基因植物的研究 近几年来,人们对植物病毒的作用机制研究逐渐明确,主要是针对病毒外壳在传播和侵染过程中作用的研究。主要集中在:利用植株自身抗病毒基因的研究[39-43];针对病毒外壳蛋白的抗病毒的植株的培育[44-46];利用基因沉默技术培育抗病植株研究[47];马铃薯病毒作为一种新型载体在基因工程中的应用研究[48]。
3 总结与展望
随着对马铃薯病毒基因组结构和功能的进一步深入研究,越来越多病毒的基因组测序完成,而且对基因功能的研究也不断利用转基因植物予以证实。世界各国已成功克隆出用于转化马铃薯的病毒外壳蛋白基因、复制酶基因、蛋白酶基因、基因调控区序列、核酶cDNA以及其他各种基因约20余种;建立完善了致瘤农杆菌介导的马铃薯转化技术;通过外壳蛋白介导、复制酶基因介导、表达基因调控区等多种途径,获得抗PVX、抗PVY、抗PVX和PVY、抗PVY和PLRV、抗PLRV、抗PSTV的转基因马铃薯栽培种。同时对病毒的生活史、检测技术、复制与表达、传播机制又有了更清楚的认识,为有效地控制马铃薯病毒的发生和危害提供了理论依据。但仍有许多方面的问题有待深入研究,主要集中在:基因组的翻译机制、后加工修饰及蛋白-蛋白作用的研究相对薄弱;与病毒结合的结构域的鉴定;病毒与宿主基因水平的互作研究,病毒基因编码蛋白在病毒传播中的研究;病毒侵染的调控机制研究等。
基于上述问题,植物病毒基因功能组学、蛋白质组学以及代谢组学的研究逐渐成为未来病毒研究的主要方向。目前蛋白质组学及代谢组学相关技术用于病毒研究主要集中在动物病毒方面的研究[49-50],为植物病毒的研究奠定了基础。因此,借助高通量、高分辨率的基因、蛋白功能分析技术,利用生物信息学分析手段,必将加快对植物病毒基因功能及蛋白结构域的分析,筛选与复制和感染有关的调控因子,有助于更深层面地了解植物病毒的生物代谢及与宿主的互作关系,病毒的生活周期等。从而为解决人类赖以生存的粮食作物病毒的侵害提供了新的途径,有助于粮食作物增产,缓解世界粮食危机。
[1]Grammatikaki G,Avgelis A.Behavior of Potato Gametoclonal Plants Against the Necrotic Strain of Potato Y Potyvirus[J].Russian Journal of Plant Physiology,2007,54(4):507-512.
[2]Makkouk K,Kumari S.Molecular Diagnosis of Plant Viruses[J].Arab J Pl Prot,2006,24(2):135-138.
[3]吴凌娟,张雅奎.用指示植物分离鉴定马铃薯轻花叶病毒(PVX)的技术[J].中国马铃薯,2003,17(2):82-83.
[4]吕典秋,李学湛,何云霞,等.烟草花叶病毒(TMV)和马铃薯X病毒(PVX)的提纯及抗血清的制备[J].中国马铃薯,2000,14(4):212-215.
[5]Khurana S M P.Pathological Problems of Economic Crop Plants and Their Management[J].Jodhpur(India):Scientific Publishers,1998:656.
[6]Corbett M K.Detection of viruses and diagnosis of plant viral diseases by electron microscopyishs,international symposium on virus diseases of ornamental plants[J].Acta Horticulturae,1972,36:III.
[7]李之芳.中国马铃薯主要病毒图鉴[M].北京:中国农业出版社,2004,5:24-69.
[8]Webster C G,Wylie S J,Craig G,et al.Diagnosis of plant viral pathogens[J].Current Science,2004,86(12):1604-1607.
[9]郭兴启,李向东.马铃薯Y病毒两分离物特性及其侵染寄主的超微结构比较研究[J].浙江大学学报:农业与生命科学版,2002,28(1):411-4l6.
[10]周艳玲,刘学敏,孟玉芹.马铃薯Y病毒的检测技术[J].中国马铃薯,2000,14(2):89-93.
[11]Singh R P,Mc Laren D L,NIE X,et al.Possible Escape of a Recombinant Isolate of Potato virus Y by Serological Indexing and Methods of its Detection[J].Plant Disease,2003:679-685.
[12]刘卫平.快速ELISA法鉴定马铃薯病毒[J].马铃薯杂志,1997,11(1):11-12.
[13]白艳菊,李学湛.应用DAS-ELISA法同时检测多种马铃薯病毒[J].中国马铃薯,2000(3):143-144.
[14]张国柱.马铃薯病毒检测技术NCM-ELISA的应用[J].山西农业科学,1991(4):25-26.
[15]孟清,宋伯符,张鹤龄.应用Dot-ELISA检测PVX、PVY和 PVS[J].中国病毒学,1993,8(4):355-371.
[16]Barker H,McGeachy K D,Toplak N,et al.Comparison of Genome Sequence of PVY Isolates with Biological Properties[J].Am J Pot Res,2009,86:227-238.
[17]Aramburu J,Galipienso L,Matas M.Characterization of potato virus Y isolates from tomato crops in northeast Spain[J].European Journal of Plant Pathology,2006,115:247-258.
[18]Abdullahi I,Koerbler M,Winte S,et al.The 18S rDNA sequence ofSynchytrium endobioticum and its utility in microarrays for the simultaneous detection of fungal and viral pathogens of potato[J].Appl Microbiol Biotechnol,2005,68:368-375.
[19]Pastrik K H.Detection of Clavibactermichiganensis subsp.sepedonicus in potato tubersbymultiplex PCR with coamplification of host DNA[J].European Journal of Plant Pathology,2000,106:155-165.
[20]Glais L,Kerlan C,Tribodet M,et al.Molecular characterization of potato virus yN isolates by PCR-RFLP Differentiation of PVYN isolates by PCR-RFLP[J].European Journal of Plant Pathology,1996,102:655-662.
[21]叶明,盛长忠,石明,等.单酶法RT-PCR检测马铃薯纺锤块茎类病毒[J].华北农学报,2002,17(1):41-45.
[22]Weidemann H L,Buchta U.A simple and rapid method for the detection of potato spindle tuber viroid(PSTVd)by RT-PCR[J].Potato Research,1998,41(1):1-8.
[23]Liu H Q,Fu Y P,Jiang D H,et al.A Novel Mycovirus That Is Related to the Human Pathogen Hepatitis E Virus and Rubi-Like Viruses[J].Journal of Virology,2009,83(4):1981-1991.
[24]Kekarainen T,Savilahti H,ValkonenJP T.Functional Genomics on Potato Virus A:Virus Genome-Wide Map of Sites Essentia for Virus Propagation.Genome Research[M].Cold Spring Harbor Laboratory Press,ISSN 1088-9051/01,2002:584-594.
[25]Kendall A,McDonald M,Bian W,et al.Structure of Flexible Filamentous Plant Viruses[J].Journal of Virology,2008,82(19):9546-9554.
[26]Zhong X H,Leontis N,Qian S M,et al.Tertiary Structural and Functional Analyses of a Viroid RNA Motif by Isostericity Matrix and Mutagenesis Reveal Its Essential Role in Replication[J].Journal of Virology,2006,80(17):8566-8581.
[27]Guenther R H,Sit T L,Gracz H S,et al.Structural characterization of an intermolecular RNA-RNA interaction involved in the transcription regulation element of a bipartite plant virus[J].Nucleic Acids Research,2004,32(9):2819-2828.
[28]Miller W A,White K A.Long-Distance RNA-RNA Interactions in Plant Virus Gene Expression and Replication[J].NIH-PA Author Manuscript Annu Rev Phytopathol,2006,44:447-467.
[29]HU Bin,PILLAI-NAIR N,HEMENWAY C.Long-distance RNA-RNA interactions between terminal elements and the same subset of internal elements on the potato virus X genome mediate minus-and plus-strand RNA synthesis[J].RNA,2007,13:267-280.
[30]Korneeva V S,Cameron C E.Structure-Function Relationships of the Viral RNA-dependent RNA Polymerase:fidelity,replication speed,and initiation mechanism determined by a residue in the ribose-binding pocket[J].Manuscrip J Biol Chem,2007,282(22):16135-16145.
[31]Plchova H,Cerovska N,Moravec T,et al.Short communication:Molecular analysis of Potato leafroll virus isolates from the Czech Republic[J].Virus Genes,2009,39:153-155.
[32]StuartG,MoffettK,Bozarth R F.A wholegenome perspective on the phylogeny ofthe plant virus family Tombusviridae[J].Arch Virol,2004,149:1595-1610.
[33]Betancourt M,Fereres A,Fraile A,et al.Estimation of the Effective Number of Founders That Initiate an Infection after Aphid Transmission of a Multipartite Plant Virus[J].Journal of Virology,2008,82(24):12416-12421.
[34]Ivanov K I,Puustinen P,Gabrenaite R,et al.Phosphorylation of the Potyvirus Capsid Protein by Protein Kinase CK2 and Its Relevance for Virus Infection[J].The Plant Cell,2003,15:2124-2139.
[35]Yelina N E,Savenkov E I,Solovyev A G,et al.Long-Distance Movement,Virulence,and RNA Silencing Suppression Controlled by a Single Protein in Hordei-and Potyviruses:Complementary Functions between Virus Families[J].Journal of Virology,2002,76(24):12981-12991.
[36]Fedorkin O N,Solovyev A G,Yelina N E,et al.Cell-to-cell movement of potato virus X involves distinct functions of the coat protein[J].Journal of General Virology,2001,82:449-458.
[37]Brault V,Bergdoll M,Mutterer J,et al.Effects of Point Mutations in the Major Capsid Protein of Beet Western Yellows Virus on Capsid Formation,Virus Accumulation,and Aphid Transmission[J].Journal of Virology,2003,77(5):3247-3256.
[38]Hofius D,Maier A T,Dietrich C,et al.Capsid Protein-Mediated Recruitment of Host DnaJ-Like Proteins Is Required for Potato Virus Y Infection in Tobacco Plants[J].Journal of Virology,2007,81(21):11870-11880.
[40]Tameling W I L,Baulcombe D C.Physical Association of the NB-LRR Resistance Protein Rx with a Ran GTPase-Activating Protein Is Required for Extreme Resistance to Potato virus X[J].The Plant Cell,2007,19:1682-1694.
[41]Kreft O,Hoefgen R,Hesse H.Functional Analysis of Cystathionineγ-Synthase in Genetically Engineered Potato Plants[J].Plant Physiology,2003,131:1843-1854.
[42]Riewe D,Grosman L,Fernie A R,et al.The Potato-Specific Apyrase Is Apoplastically Localized and Has Infiuence on Gene Expression,Growth and Development[J].Plant Physiology,2008,147:1092-1109.
[43]Sato M,Nishikawa K.Potato virus Y resistance gene,Rychc,mapped to the distal end of potato chromosome 9[J].Euphytica,2006,149:367-372.
[44]王春香,杨美珠,潘乃穟,等.马铃薯X病毒外壳蛋白基因在转基因烟草植株中的表达及抗病[J].植物学报,1993,35(11):819-824.
[45]项瑜,杨兰英,周雪荣,等.表达马铃薯Y病毒外壳蛋白基因的转基因烟草的抗病性研究[J].病毒学报,1995,11(2):158-162.
[46]宋艳茹,马庆虎,候林林,等.转PVY外壳蛋白基因马铃薯及其田间实验[J].植物学报,1996,38(9):711-718.
[47]Wang M B,Bian X Y,Wu L M,et al.On the role of RNA silencing in the pathogenicity and evolution of viroids and viral satellites[J].PNAS,2004,101(9):3275-3280.
[48]Donini M,Lico C,Baschieri S,et al.Production of an Engineered Killer Peptide in Nicotiana benthamiana by Using a Potato virus X Expression System[J].Applied and Environmental Microbiology,2005,71(10):6360-6367.
[49]常锐,刘畅,乔文涛.蛋白质组学技术在病毒学研究中的应用[J].病毒学报,2008,24(3):239-242.
[50]朱勇飞,张天宝.代谢组学在毒理学研究中的应用[J].国外医学卫生学分册,2005,32(3):156-159.
Research advances of potato virus
MA Xue-qing,WANG Yong-gang,ZHOU Xian-jing,ZHAO Hu-biao,LI Kun-peng,MA Jian-zhong*
(College of Life Science and Engineering,Lanzhou University of Technology,Lanzhou 730050,China)
Q939.46
A
1002-0306(2010)10-0429-06
2009-10-12 *通讯联系人
马雪青(1983-),在读硕士,从事植物分子生物学研究。
猜你喜欢
红蜻蜓·低年级(2022年5期)2022-05-11
当代水产(2022年1期)2022-04-26
少儿科学周刊·儿童版(2021年21期)2021-12-11
幼儿园(2021年16期)2021-12-06
今日农业(2021年4期)2021-06-09
红领巾·探索(2020年5期)2020-05-19
家教世界·创新阅读(2020年12期)2020-01-11
陕西画报(2016年1期)2016-12-01
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
