
巴马香猪Toll样受体4基因cDNA的克隆及生物信息学分析
2010-09-09 09:20巨向红徐汉进雍艳红安立龙效梅许英梅
中国实验动物学报 2010年3期
巨向红,徐汉进,雍艳红,安立龙,效梅,许英梅
(1.广东海洋大学动物医学系,湛江 524088;2.广东海洋大学动物科学系,湛江 524088)
研究报告
巴马香猪Toll样受体4基因cDNA的克隆及生物信息学分析
巨向红1,徐汉进2,雍艳红1,安立龙2,效梅1,许英梅2
(1.广东海洋大学动物医学系,湛江 524088;2.广东海洋大学动物科学系,湛江 524088)
目的研究TLR4在猪自然免疫中的作用及机制,为抗病育种及免疫佐剂的开发提供依据。方法利用NCBI公布的TLR4基因序列设计引物,RT-PCR技术克隆巴马香猪TLR4基因。结果所得基因序列提交GenBank,登录号:GQ304754。经序列分析,发现巴马香猪TLR4基因开放阅读框长2526 bp,编码785个氨基酸,该蛋白等电点为6.58,分子量为96.4×103;与普通猪比对发现巴马香猪TLR4基因有5个碱基发生突变;与小鼠、狗、鸡、牛、羊和人的同源性分别为71.9%、81.5%、54.2%、86.4%、85.5%和81.9%;TLR4膜外区蛋白为背侧多个α螺旋和内侧多个β折叠平行交替排列构成一个弯曲状螺旋结构;N末端存在信号肽,且可能在23~24位氨基酸处存在裂解位点;胞外区有13个明显的LRR,分别位于第53~74、77~100、101~124、149~173、174~192、201~225、372~393、398~429、446~469、470~494、495~518、519~541、543~566位氨基酸区;膜外区含8个N连接的糖基化位点。结论本研究成功克隆巴马香猪TLR4基因,为进一步研究该基因的功能和蛋白质的特性奠定了基础。
猪;Toll样受体;生物信息学分析
Toll-like receptors 4(TLR4)表达于许多的免疫和非免疫细胞,活化TLR4将诱导产生一系列的炎症介质包括细胞因子、趋化因子等从而产生强有力的炎症反应。最初的研究发现TLR4是革兰阴性细胞细胞壁成分脂多糖(LPS)的识别受体。近来的研究显示紫杉醇、呼吸道合胞病毒的融合蛋白、鼠乳腺肿瘤病毒的包膜蛋白、热休克蛋白60(heat shock protein 60,HSP60)、HSP70,纤连蛋白透明质酸寡聚糖、硫酸肝素多聚糖片段、纤维蛋白原以及HMGB1 ju均能活化TLR4[1,2],表明TLR4在抗细菌、抗病毒的炎症反应及应激状态下均发挥重要作用,深入研究TLR4意义重大。
巴马香猪产于广西巴马县及周边地区,其肉味鲜香,乳猪可食并鲜嫩可口,故得名“香猪”[3]。由于处在交通闭塞的边远山区,经过长期的近交,基因高度纯合,并且其体型矮小,性成熟早,已成为生命科学研究中重要的模式动物。但目前尚未发现有关巴马香猪TLR4的研究,因此本研究通过RT-PCR克隆了巴马香猪TLR4基因,通过生物信息学方法对其进行结构分析,为进一步研究猪TLR4基因的功能及其在免疫调节中的作用奠定基础。
1 材料与方法
1.1 实验样品
前腔静脉采集12头2月龄健康巴马香猪血液。淋巴细胞分离液分离外周血单个核细胞。巴马香猪购自广西巴马香猪原种场。
1.2 主要试剂
RNAiso plus、pMD18-T载体、PrimeScript 1st strand cDNA synthesis kit、Agarose gel DNA purification kit、ExTaq酶购自大连TaKaRa公司,淋巴细胞分离液购自广州展晨公司。大肠杆菌工程菌DH5α为本实验室保存。
1.3 方法
1.3.1 引物设计与合成:根据GenBank已发表的猪TLR4基因序列(NM_001113039),利用Prem ier 5.0软件设计扩增TLR4基因编码区全长PCR引物。上游引物:5′-ATGATTCCTCGCATCCGC-3′,下游引物:5′-CAAGGGACACGTTGGGAGTT-3′,扩增片段长2554 bp。引物由上海基康生物公司合成。
1.3.2 RT-PCR:首先提取巴马香猪外周血单核细胞总RNA,具体步骤按照RNAiso plus试剂盒说明书进行。总RNA用DEPC水充分溶解,琼脂糖凝胶电泳鉴定RNA完整性,-70℃保存备用。根据PrimeScript反转录试剂盒进行cDNA第一链合成,具体步骤按说明书进行。然后PCR扩增,反应体系为:14.3 μL灭菌二蒸水,2.5 μL 10×PCR buffer,2.5 μL 2.5 mmol/L dNTP Mix,1 μL 10 μmol/L primer I,1 μL 10 μmol/L primer II,0.2 μL 0.5 U/ μL Taq酶,1 μL cDNA模板。反应条件为94℃5 min,94℃1 min,55℃30 s,72℃2 min,30个循环。扩增产物经琼脂糖凝胶电泳检测。
1.3.3 目的基因回收与测序:经琼脂糖凝胶电泳鉴定有特异性条带,回收纯化产物,连接于pMD-18T载体,转化感受态DH5α菌,涂布平板,进行蓝白斑筛选,挑取阳性克隆接种于10 m L LB培养基中,过夜培养后抽提质粒,经PCR扩增进行初步鉴定,阳性质粒送上海英俊生物技术有限公司测序。
1.3.4 序列分析与结构预测:利用DNAman软件将克隆所得到的序列与GenBank上已发表的普通猪、人、小鼠、牛、羊和鸡的同源性进行比较;用NCBI中的ORF Finder软件进行开放阅读框的识别;翻译成氨基酸序列,ESyPred3D Web Server 1.0同源建模对TLR4蛋白进行三维结构预测;Signal P3.0预测TLR4蛋白的N端信号肽结构,TMHMM Server v. 2.0预测跨膜结构,SMART service预测保守结构域,NetGlyc1.0 Server进行N-糖基化位点预测。
2 结果
2.1 RNA完整性及PCR产物的特异性分析
对提取的RNA进行电泳分析,结果可明显地观察到28 s、18 s和5 s三条带(图1-A),说明所提取RNA完整性良好。对RT-PCR产物的特异性进行电泳分析,结果显示TLR4 PCR产物特异性良好,在2500 bp左右出现目的条带(2554 bp)(图1-B)。
2.2 重组质粒PCR鉴定
质粒电泳结果显示2500 bp左右有目的条带(2554 bp)出现(图1-C),说明TLR4目的片段已插入到载体当中。
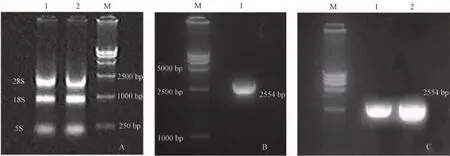
注:A为RNA电泳结果;1、2为RNA样品,M为分子质量标准。B为PCR产物电泳结果;M为分子质量标准,1为PCR产物。C为重组质粒PCR产物电泳图;M为分子质量标准,1、2为重组质粒PCR产物。图1 RNA、PCR产物及重组质粒电泳结果Note:Figure 1A shows the electrophoresis result of total RNA.1 and 2 represent RNA sample and M is marker.Figure 1B shows the electrophoresis results of RT-PCR products of TLR4 gene.M is marker and 1 is the RT-PCR product.Figure 1C shows the electrophoresis result of recombinant p lasmid PCR products.M is marker and 1 and 2 are the recombinant plasmid PCR products.Fig.1 The results of electrophoresis of Total RNA,RT-PCR products of TLR4 and recombinant plasmid.
2.3 TLR4基因测序结果及分析
核苷酸序列测序结果表明,巴马香猪TLR4基因开放阅读框全长2358 bp,编码785个氨基酸。将巴马香猪TLR4测序结果用DNAman软件与GenBank中登录的猪TLR4(NM_001113039)的核苷酸序列及氨基酸序列比对,发现巴马香猪TLR4编码区核苷酸序列有5个碱基被替换,结果造成了5个无义突变(表1)。突变位置在图6中以粗线方框标出。
2.4 巴马香猪TLR4的同源性分析
用DNAman软件将巴马香猪TLR4测序核苷酸序列与GenBank中登录的普通猪(NM213761)、狗(NM001005264)、鸡(NM204278)、牛(NM174197)、人(NM003264)、小鼠(NM011905)和羊(NM001048231)的参考序列进行比对(图2),发现巴马香猪与普通猪同源性达到99.8%,与牛和羊的同源性较高,分别为86.4%和85.5%。与人、狗和小鼠的同源性分别为81.9%、81.5%和71.9%。与鸡同源性最低,仅为54.2%。将巴马香猪TLR4基因翻译成氨基酸,与GenBank中登录的普通猪、狗、鸡、牛、人、小鼠、羊参考序列进行比对,发现同源性分别达到99.4%、70.8%、44.9%、80.0%、72.5%、62.4%和79.2%。预测巴马香猪TLR4蛋白等电点为为6.58,分子量为96.4×103。
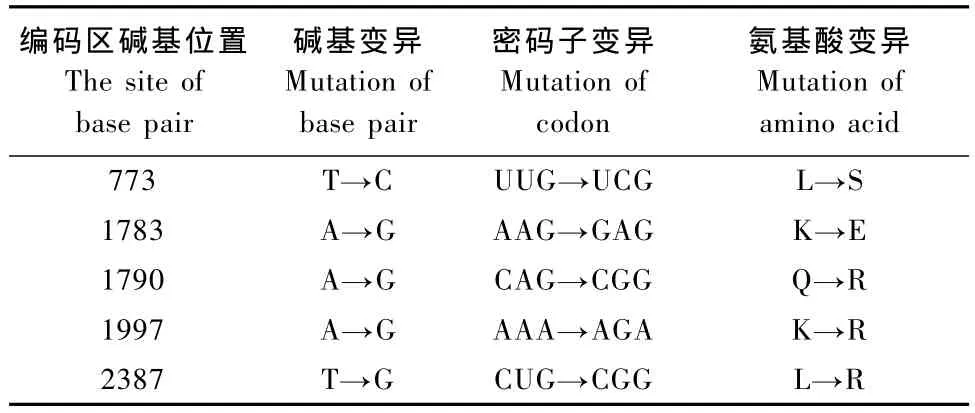
表1 与普通猪进行序列比对巴马香猪TLR4碱基突变Tab.1 The TLR4 base mutation in Bama miniature pig compared with that in common pigs

注:“-”表示保守氨基酸;“.”表示氨基酸缺失。图2巴马香猪TLR4氨基酸序列与其他动物比对Note:“-”indicates conservative amino acid;“.”indicates amino acid deletion.Fig.2 Comparison of the identical alignment of the amino acid sequences of Bama miniature pig TLR4 with that of common pig,cattle and chicken.
2.5 三维结构预测
ESyPred3D Web Server 1.0同源建模对TLR4蛋白进行三维结构预测,得到TLR4膜外区的立体三维结构。预测结果表明,TLR4膜外区蛋白由背面多个α螺旋和内面多个β折叠构成。α螺旋和β折叠平行交替排列,构成一个弯曲状螺旋结构(图3,彩插1)。
2.6 TLR4信号肽预测
Signal P3.0预测TLR4蛋白的N端信号肽结构,其中Signal P-NN预测信号肽裂解点[4]。结果表明,在23~24位氨基酸可能存在裂解位点。
2.7 跨膜结构分析
TMHMM Server v.2.0预测跨膜结构。结果表明,巴马香猪TLR4分子可能是由胞外区(1-588)、跨膜区(589-611)、胞内区(612-785)3部分组成的膜蛋白(图4,彩插1)。
2.8 保守结构域分析
SMART service预测保守结构域,发现该蛋白胞外区有13个明显的LRR,分别位于第53~74,77~100,101~124,149~173,174~192,201~225,372~393,398~429,446~469,470~494,495~518,519~541,543~566位氨基酸区(图5,彩插1)。13个LRR在图2三维结构中分别以黄色标记、分别为LRR1、LRR2、LRR3、LRR4、LRR5、LRR6、LRR7、LRR8、LRR9、LRR10、LRR11、LRR12、LRR13。在LRR区之后是胞外肽链C端基序LRR-CT(579-628)。
2.9 糖基化位点分析
根据NetGlyc1.0 server进行N-糖基化位点预测,发现TLR4胞外区存在8个N-连接的糖基化位点(图6,以细线方框标记)。N-糖基化位点在图3三维结构中采用RasMol软件以紫色标记为Asn35、Asn205、Asn238、Asn282、Asn309、Asn526、Asn575和Asn625。

注:细线方框标记为预测的糖基化位点,粗线框方框标记为氨基酸突变位点。图6 TLR4蛋白糖基化位点预测Note:The glycosylation sites are marked with fine frame and mutation of amino acid with coarse frameFig.6 The predicted glycosylation sites of the TLR4 protein
3 讨论
随着对TLR4的分子结构、识别受体、信号传导途径及基因缺陷型动物模型等研究的深入,其在机体免疫系统中的重要性也日益得以认识。本实验运用RT-PCR方法克隆了巴马香猪TLR4基因编码区,与普通猪TLR4基因进行序列比对,巴马香猪TLR4出现了5处有义突变,其中两个突变发生在胞内区。由于胞内区与信号的传导密切相关,因此胞内区的突变很容易影响TLR4正常功能的发挥以及对疾病的易感性。White等[5]发现牛TLR4编码区第274~368残基位置是一个变异频发区,这一区域在不同物种之间保守性非常低,即使在同一物种内突变率也较高,因此认为此位置参与对病原分子的连接绑定。Thomas等[6]认为这一变异频发区的多态性可能反映了猪对疾病易感性模式的不同。比对发现巴马香猪TLR4与普通猪TLR4在这一变异频发区序列完全一致,没有发生任何变异突变。对其核苷酸序列的同源性比较后发现,巴马香猪TLR4与普通猪同源性极高,与牛、羊、狗、人和鼠有较高相似性,与鸡同源性较差。这说明在TLR4长期的生物进化过程中具有较强保守性。
同源建模预测分析表明,巴马香猪TLR4膜外区蛋白由背面多个α螺旋和内面多个β折叠构成。信号肽预测表明TLR4存在典型的信号肽结构,跨膜结构预测结果表明TLR4存在典型的跨膜结构,这与Thomas等[6]报道的TLR4结构一致。Bella等[7]认为LRR结构作为蛋白质与蛋白质之间相互作用的调节模式非常适合。在目前已知的LRR绑定复合物中,一般认为LRR螺线管结构的凹面就是链接绑定位点。但最近的研究发现一些LRR结构使用他们的凹面形成稳定的二聚体[8,9],这说明LRR能用其他的绑定模式[10]。Alvarez[11]认为普通猪TLR4第279~373位置还存在4个首尾相连的串联LRR,并认为此处可能是连接绑定位点。这4个LRR在不同物种之间的同源性非常低,并且有研究认为正是TLR4的这个串联LRR区域导致了鼠和人类在区分不同类型LPS上能力有所差别[12,13]。Kobe[14]研究发现TLR5与鞭毛蛋白的连接绑定位点位于LRR14上,LRR14有32个残基,这揭示LRR14第15位后面插入了6个残基,正是这6个残基使得TLR5与鞭毛蛋白相结合。巴马香猪TLR4的胞外结构域具有13个明显的LRR,符合TLR的胞外结构特征。其中的LRR部分构成配体结合区。TLR4胞外区三维结构呈弯曲的螺线管状,其凹面LRR区富含疏水氨基酸残基,从而形成一个疏水核心。LRR4,LRR6和LRR10第10位后均有一个残基插入,而LRR1、LRR5、LRR7、LRR8和LRR12均有残基丢失,我们认为这种现象可能与TLR4功能的特异性有关。糖基化是蛋白质的一种修饰方式。主要是修饰天冬酰胺上的N,其氨基酸的特征序列是Asn-X-Ser/Thr(X代表任何一种氨基酸)。近年来发现,糖蛋白中的糖链参与细胞识别与分子识别,因此称糖蛋白中的糖链为信息分子[15]。朱立平[16]认为糖链是继核酸链与蛋白质链之后,学者们深入研究与生命活动息息相关的第三链。本研究发现TLR4胞外区存在8个N-连接的糖基化位点。其中第4和第5个糖基化位点Asn238和Asn282位于4个串联LRR上,由于此区域可能是连接绑定位点,该区域的糖基化很容易阻止TLR4与PAMPs的绑定结合。这些位置的糖基化可能与TLR4功能的发挥密切相关。
(本文图3~5见彩插1。)
[1]Akira S,Uematsu S,Takeuchi O.Pathogen recognition and innate immunity[J].Cell,2006,124(4):783-801.
[2]Van Beijnum JR,Buurman WA,Griffioen AW.Convergence and amplification of toll-like receptor(TLR)and receptor for advanced glycation end products(RAGE)signaling pathways via high mobility group B1(HMGB1)[J].Angiogenesis,2008.11 (1):91-99.
[3]王爱德,兰干球,郭亚芬.巴马香猪耐热性的探讨[J].家畜生态,1995.16(4):18-21.
[4]Bendtsen JD,Nielsen H,Heijne J,et al.,Improved prediction of signal peptides:Signal P 3.0[J].J Mol Biol,2004.340:783-795.
[5]White SN,Taylor KH,Abbey CA,et al.Haplotype variation in bovine Toll-like receptor 4 and computational prediction of a positively selected ligand-binding domain[J].Proc Natl Acad Sci U S A,2003.100:10364-10369.
[6]Thomas AV,Broers AD,Vandegaart HF,et al.Genomic structure,promoter analysis and expression of the porcine(Sus scrofa)TLR4 gene[J].Molec Immunol,2006.43:653-659.
[7]Bella J,Hindle KL,McEwan PA,et al.The leucine-rich repeat structure[J].Cell Mol Life Sci,2008.65:2307-2333.
[8]Scott PG,Dodd CM,Bergmann EM,et al.Crystal structure of the biglycan dimer and evidence that dimerization is essential for folding and stability of class I small leucine-rich repeat proteoglycans[J].J Biol Chem,2006.281:13324-13332.
[9]Scott PG,McEwan PA,Dodd CM,et al.,Crystal structure of the dimeric protein core of decorin,the archetypal small leucinerich repeat proteoglycan[J].Proc Natl Acad Sci U S A,2004. 101:15633-15638.
[10]McEwan PA,Scott PG,Bishop PN,et al.Structural correlations in the family of small leucine-rich repeat proteins and proteoglycans[J].J Struct Biol,2006.155:294-305.
[11]Alvarez B,Revilla C,Chamorro S,et al.Molecular cloning,characterization and tissue expression of porcine Toll-like receptor 4[J].Dev Comp Immunol,2006.30:345-355.
[12]Bell JK,Mullen GED,Leifer CA,et al.,Leucine-rich repeats and pathogen recognition in Toll-like receptors[J].Trends Immunol,2003.24:528-533.
[13]Hajjar AM,Ernst RK,Tsai JH,et al.Human Toll-like receptor 4 recognizes host-specific LPS modifications[J].Nat Immunol,2002.3:354-359.
[14]Kobe B,Deisenhofer J.Proteins with leucine-rich repeats[J]. Curr Opin Struct Biol,1995.5:409-416.
[15]Sharon H,Lis H.Carbohydratas in cell recognition[J].Sci Am,1993.268:82.
[16]朱立平.蛋白质糖基化与B细胞免疫[J].现代免疫学,2001.21(4):193-194.
Cloning and Bioinform atics Analysis of TLR4 cDNA in Bam a M iniature Pig
JU Xiang-hong1,XU Han-jin2,YONG Yan-hong1,AN Li-long2,XIAO Mei1,XU Ying-mei2
(1.Department of veterinary medicine,2.Department of Animal Science,Guangdong Ocean University,Zhanjiang 524088,China)
ObjectiveBama Miniature pig is originated in the Chengguan area of Bama County and Yiwei area of Dongtian County in Guangxi Zhuang Autonomous Region of China.The form of this breed is the results of long time inbreeding,so the heredity and production performance is very stabilized,and are used as an experiment animal model normally.Toll-like receptor 4(TLR4)is an important member of Toll-like receptor family,it mainly responses to lipopolysaccharide(LPS)and plays a vital role in the host immune system.In order to explore the molecular mechanism of TLR4 participating in immunosuppression induced by heat stress in Bama Miniature Pigs.The present study is conducted to clone the TLR4 gene cDNA and make bioinformatics analysis of the protein after translated into amino acids.M ethods Total 12 bama miniature pigs aged approximately 3-4 months were used in this study.One milliliter blood was collected into vacutainers by anterior vena cava puncture.Lymphocytes were isolated from 1 mL of whole blood by density gradient centrifugation using LTS-1077 lymphocyte separation solution.TLR4 gene was cloned from peripheral blood mononuclear cells by reverse transcription polymerase chain reaction(RT-PCR).The Identical alignment of the amino acid sequence was compared with other animals by DNAman software.Homology modeling to prediction the three-dimensional structure models of TLR4 extracellular region was conducted by ESyPred3D Web Server 1.0 software.The N-terminal signal peptide structure was predicted by Signal P3.0 software.The transmembrane region structure was predicted by TMHMM Server v.2.0 software.The conserved domain region was predicted by SMART service software and the N-glycosylation site by NetGlyc1.0 Server software.ResultsThe results showed that Toll-like receptor 4 gene consisted of 2358 base pairs.The gene sequence was submitted to GenBank(gi:GQ304754).The TLR4 gene coded 785 amino acids.The molecular weight of TLR4 protein was 96.4 kD and the isoelectric point was 6.58.This protein had a transmembrane region(589-611)and a signal peptide in N-terminal,and might be a schizolysis site in 23-24 amino acids.5 bases of TLR4 gene in Bama miniature pig were mutated compared with that of common pig,but the structure did not change significantly.The Bama miniature pig TLR4 gene sequence had a high homology compared with that of mice(73.4%),dog(82.4%),chicken(59.5%),cattle(85.3%),sheep(84.6%)and man(82.4%).The structure of extracellular region of TLR4 protein showed a forniciform helix structure,and consisted of a lot of α-helix in inside and β-sheet in outside of the arc and they arranged parallely and alternately.There were thirteen leucine-rich repeats located at 53-74,77-100,101-124,149-173,174-192,201-225,372-393,398-429,446-469,470-494,495-518,519-541 and 543-566 amino acid site,respectively,and there were also 8 N-linked glycosylation sites out of the membrane region.ConlusionTLR4 gene of Bama miniature pigs has been successfully cloned and the possible structure and function are pretested by bioinformatic analysis in the present study.
Pig;Toll-like receptor;Bioinformatics analysis
R349.64
A
1005-4847(2010)03-0185-06
2009-12-03
广东海洋大学博士启动基金(编号:0712107)。
巨向红(1977-),男,研究方向:动物生殖生理与免疫。E-mail:juxianghong@yahoo.com.cn;徐汉进(1982-),男,研究方向:动物营养与饲料科学专业。E-mail:pixingdy@hotmail.com,与第一作者有同等贡献。
雍艳红。Yongyanhong-007@163.com
猜你喜欢
生物技术通报(2021年6期)2021-08-11
癌症进展(2018年11期)2018-12-30
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
医学研究杂志(2015年7期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
中国粮油学报(2014年7期)2014-02-06
中国饲料(2013年1期)2013-01-25
