
芦芽山阴坡华北落叶松—云杉天然次生林群落特征的海拔梯度格局1)
2010-08-09 11:07武秀娟常建国于吉祥郭建荣
东北林业大学学报 2010年11期
武秀娟 常建国 于吉祥 安 雁 郭建荣
(山西省林学会,太原,030012) (中国林业科学研究院华北林业研究所) (山西省林学会) (山西大学) (芦芽山国家级自然保护区)
植物群落是植物与植物之间,植物与环境之间形成具有一定相互关系的植物种类的集合体[1]。生物多样性沿环境梯度的变化趋势是生物多样性研究的一个重要议题[2],物种多样性则是生物多样性的本质内容,其分布格局主要与气候、群落生产力等相关[3-4]。海拔影响着山地物种分布的非生物条件如气候和土壤等,因此,是影响山地物种组成和群落结构的重要因素[5-6],物种多样性具有多种海拔梯度分布格局,其中最普遍的就是物种多样性随着海拔的升高而不断减少[7-9],单峰分布也是一种比较常见的物种多样性随海拔梯度分布的格局[10],也有研究[11]发现物种多样性随海拔升高而增加。不同的物种其分布区的主要控制因子不同,多样性的海拔分布格局也有所差异,研究生物多样性的海拔梯度格局对于解释生物多样性的环境梯度变化规律具有重要意义[12]。
植物群落分类是植被研究中最复杂的问题之一,它不仅能使我们了解一个地区的植被特点,同时还能使我们认识相邻地区之间植被的联系规律[13]。植被数量分析在过去几十年中已成为现在植物群落生态学研究中必不可少的手段[14],近20 a来,多元分析技术在群落生态学中的应用发展很快,其中分类和排序是最为常用的植被数量分析方法。实践表明,分类和排序结合使用更能客观地反映植物群落的生态关系,是研究植被格局、植被与环境间关系的重要手段[15]。
芦芽山自然保护区主要保护寒温性森林植被类型,且该保护区是汾河的发源地,许多学者在芦芽山做过多方面的研究工作,尤其是植被生态学方面的工作[16-18],但这些研究的尺度都比较大,涉及的海拔跨度大,森林类型众多,未能对华北落叶松—云杉天然次生林小尺度群落特征的海拔格局给出定论,因此,本研究以华北落叶松—云杉天然次生林为对象,通过调查不同海拔梯度该林型群落的特征及各月份的土壤体积含水量,结合常用的物种多样性测度指标(多样性指数、均匀度指数和丰富度指数)和物种最宜生长海拔高度的计算,旨在了解:不同海拔梯度的林分特征;灌草植物物种多样性的海拔梯度格局;灌草植物分布及其与环境间的关系。
1 研究区概况
研究地位于山西省芦芽山国家级自然保护区,北纬38°36'~39°02',东经111°46'~112°54'。该区属暖温带半湿润区,具有明显的大陆性气候特点,夏季凉爽多雨,冬季寒冷干燥。多年平均气温为5℃,1月份均温-9℃,7月份均温为20℃,年降水量400 mm,年蒸发量1 800 mm,年均相对湿度55%,无霜期130 d。土壤主要是在残积和坡积母质上发育起来的。芦芽山海拔1 850 m以上地段基本上是以华北落叶松(Larixprincipis-rupprechtii)和云杉属植物[白杄(Piceameyeri)、青扦(Picea wilsonii)]为建群种组成的寒温性针叶林占优势[16]。
2 材料与方法
2.1 群落调查
华北落叶松—云杉天然次生林群落调查采取样方法,样方沿海拔梯度布设,且避免人为扰动和特殊的生境。2009年8月份在海拔2 000、2 100、2 200、2 400和2 600 m处分别设置4个25 m×25 m的乔木样地,每个样地内设5个5 m×5 m的灌木样方和5个2 m×2 m的草本样方,共计20个乔木样地,100个灌草样方。调查记录每个样地的坡度、乔木物种组成、个体高度、胸径、第一活枝下高和最大冠幅;灌草样方中分别进行物种组成,灌木和草本的高度、盖度与数量等方面的野外调查[19]。在各样地内应用时域反射仪(Time Domain Reflectometry,简称TDR)测定0~10cm的土壤体积含水量,自2009年5月份开始,每月8日、18日、28日测定,雨后加测,到9月末为止。
2.2 相关指数的计算方法
灌木和草本的数量指标用重要值度量[20],物种多样性选用物种丰富度指数、物种多样性指数(Shannon-Wiener指数[21]、Simpson 指数[22])和均匀度指数(Pielou 指数[23]、Alatalo指数[24])进行测度,物种最宜生长海拔使用各物种重要值的加权平均值表示[25],各指数计算公式如下:


式中,Pi为种i的相对重要值;H'为Shannon-Wiener指数;D为 Simpson 指数;Js,w、Js,i均为 Pielou 指数;Ea为 Alatalo 指数;S为种i所在样方的物种总数,即丰富度指数;WA为最宜生长海拔高度;H为海拔高度。
2.3 数据分析
运用SPSS16.0的One-way ANOVA模块对不同海拔梯度乔木层各参数进行方差分析,同时用最小显著差数法(LSD)进行多重比较。利用灌草植物的重要值信息在PCORD 5的双向指示种分析(TWINSPAN)模块下[“假种(Pseudospecies)”分级水平值分别是 0、0.02、0.05、0.10、0.20]进行群落分类,同时在去趋势对应分析(DCA)模块下对不同海拔梯度的样方和物种进行分析。根据植被生态学和数量生态学方法的要求,把频度≤5%的物种剔除[18],选出具有代表性的41个物种进行研究,并作分析。
3 结果与分析
3.1 不同海拔的林分特征
随海拔的升高,林分密度逐渐增加,海拔2 000 m处最小,为800株·hm-2,海拔2 600 m处最大,为1 952株·hm-2,且海拔2600 m处的林分密度明显大于其他海拔处的;林分的胸高断面积呈现出随海拔的升高在波动中增加的趋势,海拔2 000 m处的明显低于其他海拔处的;平均树高、平均胸径则表现出随海拔的升高而呈波动中减少的趋势,说明随着海拔的升高乔木层植被变细,变矮;平均活枝下高随着海拔的升高有上升的趋势,在海拔2400 m及以上海拔处的平均活枝下高明显高于低海拔处的;平均冠幅则随着海拔的升高呈减小的趋势,表明沿海拔梯度的变化林分的活冠层有变窄的倾向(表1)。
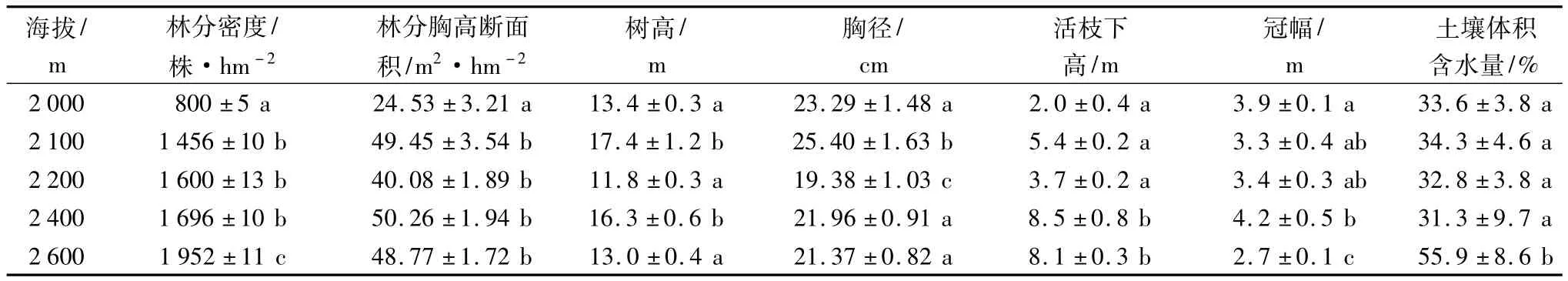
表1 不同海拔梯度林分及土壤水分特征
3.2 不同海拔灌草群落物种多样性特征
表2为不同海拔灌木和草本群落的物种多样性,可以看出,灌木和草本层的多样性指数,即Shannon-Wiener指数(H')和Simpson指数(D)的变化趋势基本一致,几乎均表现为草本层的高于灌木层的。灌木层的H'值与D值有随海拔明显减小的趋势(p<0.05),草本层的H'值与D值随海拔增加出现波动,但变化不大(p>0.05)。灌木层物种丰富度指数(S)随海拔有减小的趋势,草本层的则呈增加趋势,但两者都不明显(p>0.05),且该指数在草本层大于灌木层,可见,不同海拔灌草群落物种的丰富度和物种多样性主要由草本层决定。
灌木层的 3 个均匀度指数Js,w、Js,i和Ea随海拔的变化呈现出相似的变化趋势(表2),即随海拔变化明显减小(p<0.05);草本层的这3个值则表现为随海拔的升高,变化趋势均不明显(p>0.05)。除海拔2 100 m和2 600 m外,灌木层的均匀度指数都高于草本层,在海拔2 100 m处草本层均匀度虽高于灌木层,但差值很小,最大差值仅是其他海拔灌木层高于草本层的1/2。海拔2 600 m处则无灌木出现。

表2 不同海拔梯度灌草层植物的物种多样性
3.3 灌草群丛的类型、分布格局及与环境因子的关系
3.3.1 不同海拔的灌草群丛类型及特征
采用TWINSPAN对华北落叶松—云杉天然次生林林下灌草层的100个样方进行分类,结果分为25组,按照中国植被分类原则及结合野外实际调查的情况,对TWINSPAN的分类结果进行合并,将25个组合并成13组,代表13个林下灌草群丛,这样能更客观地反应林下各群丛的组成情况,各群丛的主要特征如表3:从表3中可看出,海拔2 000~2 200 m处主要优势灌木包括灰栒子、虎榛子、黄刺玫、金花忍冬和葱皮忍冬,海拔2 400 m处这些优势灌木全部消失,被以八宝茶为主的优势灌木所取代,海拔2600m处灌木则全部消失,由此看来,海拔2 200~2 400 m是灌木物种组成发生明显变化的位置。从调查结果推断灌木物种分布的上限在海拔2 400~2 600 m(表3),补充调查发现,其确切位置为海拔2 550 m;优势草本中仅披针苔草在海拔2000~2600 m均有分布,其他优势草本自海拔2400 m始为苔藓取代,可见,海拔2 200~2 400 m处优势草本组成也发生很大变化,披针苔草这一广泛分布优势种的发现对于研究该区域物种对环境的适应机制有重要价值。
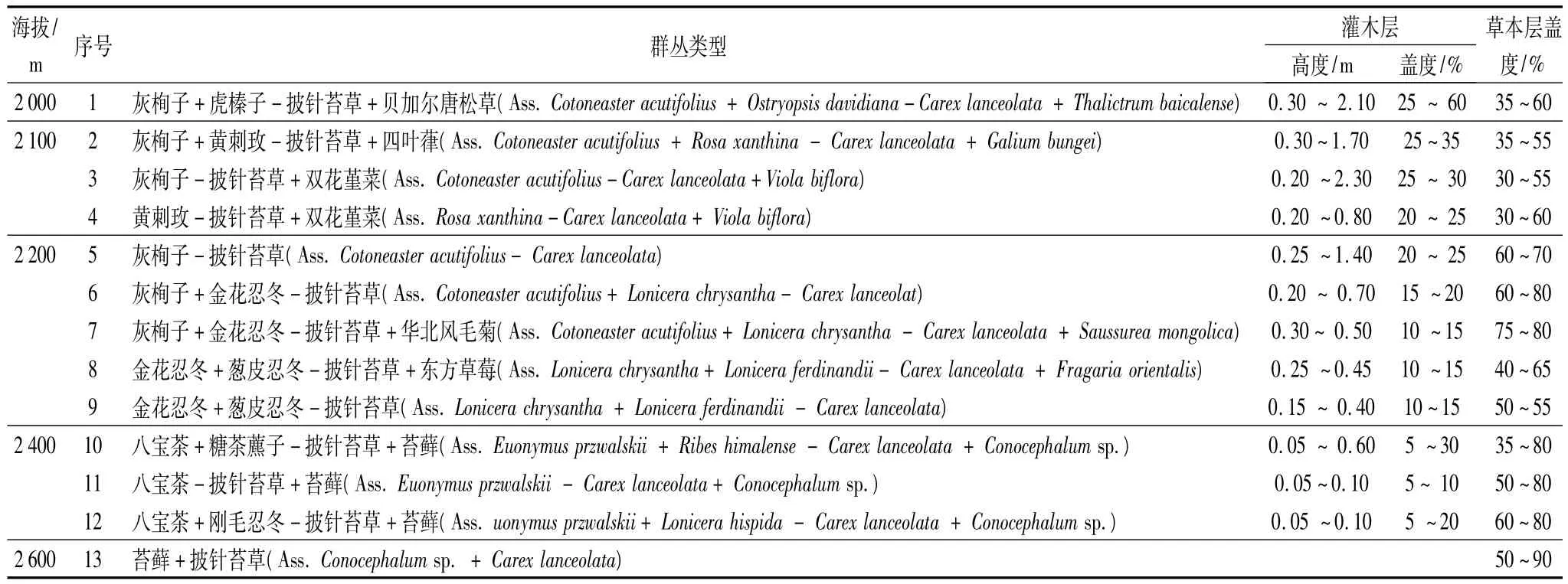
表3 不同海拔的灌草群丛类型及特征
3.3.2 灌草植物分布与环境的关系
DCA分析表明前两个轴包含的生态信息较多,显示出重要的生态意义(第一排序轴的特征值为0.737,第二排序轴的特征值为0.443)(图1)。图1是100个样方灌草植物的DCA二维排序图,TWINSPAN分类所产生的13个灌草植物群丛在图1中各有自己的分布范围和界限,说明DCA较好地反映出灌草植物之间以及与环境之间的关系。
从排序轴来看,第一排序轴基本上反映灌草植物与海拔之间的关系,从A区到D区海拔由低到高(图1)。排序图中100个样方共划分成4个主要生态区,A区主要含群丛1~4,由海拔2000 m和2 100 m处样方组成;B区主要含群丛5~9,由海拔2 200 m处样方组成;C区主要含群丛10~12,由海拔2 400 m处样方组成;D区主要含群丛13,由海拔2 600 m处样方组成。第二排序轴反映了灌草植物所在环境的水分梯度变化趋势,从下向上,水分依次增大(表1)。
图2中是41个物种DCA排序的二维图形,排序轴所反映的生态意义与图1相同,灰栒子、黄刺玫和虎榛子为海拔2000、2 100 m处的优势灌木;金花忍冬和葱皮忍冬属海拔2 200 m处的优势灌木;八宝茶属海拔2 400 m处的优势灌木。草本植物也随着第一排序轴有所变化,如图2中小红菊和贝加尔唐松草位于第一排序轴的左边,即处于低海拔处,羽节蕨和藓类位于第一排序轴的右侧,即高海拔处,同时小红菊、贝加尔唐松草属耐干旱型草本,羽节蕨、藓类属喜湿润型草本,进一步说明了环境水分含量的大小也是影响灌草植物分布的一个不可忽视的因子。
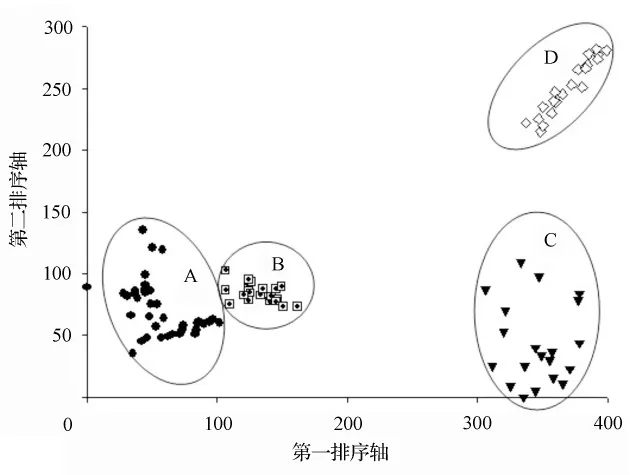
图1 100个样方的DCA二维排序

图2 41个物种的DCA二维排序

表4 灌草植物的最宜生长海拔
各灌草植物最适宜分布的海拔如表4中所示,从表4中可看出,灌草植物物种的分布与图2中第一排序轴反映的生态意义即海拔特征基本吻合,进一步说明海拔是限制林下灌草植物最优分布区域的一个重要因素。由表4还可看出在不同的海拔,适宜生长的物种数量总体上没有明显变化,但海拔2 400 m以上的灌木物种数明显减少,草本物种数明显增加。
4 结论与讨论
随海拔上升,乔木层平均树高、平均胸径在波动中下降,这与赵淑清等[26]、Gao Junfeng 等[27]的研究结果不同,但与沈泽昊等[28]的一致,这可能是在中低山与亚高山区,影响林木生长的主导环境因子组成及各因子作用力的权重存在差异。此外,林木对资源的竞争也直接影响其生长,如本研究发现随海拔升高,林分密度增加,林木竞争加剧,树高与胸径变小。因此,构建较大空间尺度上的森林生长动态模型时,至少应同步考虑环境因子与林木竞争。
与多数研究相同[9,29-32],灌木层多样性指数与均匀度指数均随海拔上升而减小,可能是环境能量(如年均温等)随海拔减小所致,符合多样性—环境能量假说。草本层多样性指数与均匀度指数沿海拔变化不大,与Ohlemueller等[33]及赵淑清等[32]的研究结论相同,方精云[34]认为草本层物种多样性主要受控于乔木层郁闭度而非海拔,本研究也发现,各海拔梯度上的乔木层郁闭度基本保持在0.70左右。
多数学者常采用TWINSPAN植被数量等级分类方法,结合当地植被情况,将“假种”划分为5~7级,并据此将植物群落划分为8~23类[35-37]。本研究采用了5级“假种”将海拔2 000~2 600 m的灌草植物划分为13类群丛,与张丽霞等[17]在该地海拔2 000~2 600 m的分类结果(11个)相近。
海拔变化主要表现为气候变化[38],本研究中DCA排序将13类灌草群丛划为4个区,排序轴生态意义与刘秀珍等[18]的一致,反映了海拔和土壤水分的梯度变化,可见,气候与土壤水分是决定该区域灌草植物海拔分布格局的主要环境因子。此外,研究发现灌草物种均有其最宜分布的海拔位置,这是植物与环境长期协同进化的结果,最宜分布位置的确定,对气候变化背景与情景下开展物种迁移方向与迁移速度的观测研究奠定了基础[39]。
[1]李日红.植物群落的特点和演替[J].中山大学学报论丛,2000,20(5):27-32.
[2]Odland A,Birks H J B.The altitudinal gradient of vascular plant richness in Aurland,western Norway[J].Ecography,1999,22(5):548-566.
[3]O'Brien E M,Field R,Whittaker R J.Climatic gradients in woody plant(tree and shrub)diversity:water-energy dynamics,residual variation and topography[J].Oikos,2000,89(3):588-600.
[4]Kratochwil A.Biodiversity in ecosystems:some principles,biodiversity in ecosystems[M].Kluwer Academic Publishers Dordrecht,1999:5-38.
[5]Austin M P,Pausas J G,Nicholls A O.Patterns of tree species richness in relation to environment in southeastern New South Wales,Australia[J].Austral Ecology,1996,21(2):154-164.
[6]Austrheim G.Plant diversity patterns in semi-natural grasslands along an elevational gradient in southern Norway[J].Plant Ecology,2002,161(2):193-205.
[7]Lieberman D,Lieberman M,Peralta R,et al.Tropical forest structure and composition on a largescale altitude gradient in Costa Rica[J].Journal of Ecology,1996,84:137-152.
[8]Givnish T J.On the cause of gradients in tropical tree diversity[J].Journal of Ecology,1999,87:193-210.
[9]郝占庆,于德永,杨晓明,等.长白山北坡植物群落α多样性及其随海拔梯度的变化[J].应用生态学报,2002,13(7):785-789.
[10]Kitayama K.An altitudianl transect study of the vegetation of Mount Kinabalu,Borneo[J].Plant Ecology,1992,10(2):149-171.
[11]Itow S.Species turnover and diversity pattern along an evergreen broad-leaved forest coenocline[J].Journal of Vegetation Science,1991,2(4):477-484.
[12]唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
[13]宋永昌.植被生态学[M].上海:华东师范大学出版社,2001:297.
[14]阳含熙,卢泽愚.植物生态学的数量分类方法[M].北京:科学出版社,1981:1-25.
[15]张峰,张金屯.我国植被数量分类和排序研究进展[J].山西大学学报:自然科学版,2000,23(3):278-282.
[16]张丽霞,张峰,上官铁梁.芦芽山植物群落的多样性研究[J].生物多样性,2000,8(4):361-369.
[17]张丽霞,张峰,上官铁梁.山西芦芽山植物群落的数量分类[J].植物学通报,2001,18(2):231-239.
[18]刘秀珍,张峰,张金屯.管涔山撂荒地植物群落演替过程中物种多样性研究[J].武汉植物学研究,2008,26(4):391-396.
[19]吴彦,刘庆,乔永康,等.亚高山针叶林不同恢复阶段群落物种多样性变化及其对土壤理化性质的影响[J].植物生态学报,2001,25(6):648-655.
[20]张金屯.植被数量生态学方法[M].北京:中国科学技术出版社,1995.
[21]马克平,刘玉明.生物群落多样性的测度方法I:α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[22]Magurran A E.Ecological diversity and its measurements[M].New Jersey:Princeton University Press,1988.
[23]Whittaker R H.Evolution and measurement of species diversity[J].Taxon,1972,21(2/3):213-251.
[24]Hurlbert SH.The nonconcept of species diversity:A critique and alternative parameters[J].Ecology,1971,52(4):577-586.
[25]LepšJ,Šmilauer P.Multivariate analysis of ecological data using CANOCO[M].Cambridge:Cambridge University Press,2003.
[26]赵淑清,方精云,朴世龙,等.大兴安岭呼中地区白卡鲁山植物群落结构及其多样性研究[J].生物多样性,2004,12(1):182-189.
[27]Gao Junfeng,Zhang Yunxiang.Distributional patterns of species diversity of main plant communities along altitudinal gradient in secondary forest region,Guandi Mountain,China[J].Jouranl of Forestry Research,2006,17(2):111-115.
[28]沈泽昊,刘增力,方精云.贡嘎山海螺沟冷杉群落物种多样性与群落结构随海拔的变化[J].生物多样性,2004,12(2):237-244.
[29]Ojeda F,Marai¨non and Airoyo J.Plant diversity patterns in the Aljibe Mountains(S.Spain):a comprehensive account[J].Biodiversity and Conservation,2000,9(9):1323-1343.
[30]陈廷贵,张金屯.山西关帝山神尾沟植物群落物种多样性与环境关系的研究I.丰富度、均匀度和物种多样性指数[J].应用与环境生物学报.2000,6(5):406-411.
[31]岳明,张林静,党高弟,等.佛坪自然保护区植物群落物种多样性与海拔的梯度关系[J].地理科学,2002,22(3):349-354.
[32]赵淑清,方精云,宗占江,等.长白山北坡植物群落组成、结构及物种多样性的垂直分布[J].生物多样性,2004,12(1):164-173.
[33]Ohlemueller R,Wilson JB.Vascular plant species richness along latitudinal and altitudinal gradients:a contribution from New Zealand temperate rainforests[J].Ecology Letters,2002,4:262-266.
[34]方精云.探索中国山地植物多样性的分布规律[J].生物多样性,2004,12(1)1-4.
[35]史作民,刘世荣,程瑞梅,等.河南宝天曼植物群落数量分类与排序[J].林业科学,2000,36(6):20-27.
[36]欧光龙,彭明春,和兆荣,等.高黎贡山北段植物群落TWINSPAN数量分类研究[J].云南植物研究,2008,30(6):679-687.
[37]胡刚,梁士楚,张忠华,等.桂林岩溶石山青冈栎群落的数量分析[J].生态学杂志,2007,26(8):1177-1181.
[38]刘鸿雁,曹艳丽,田军,等.山西五台山高山林线的植被景观[J].植物生态学报,2003,27(2):263-269.
[39]Lenoir J,Gégout J C,Marquet P A,et al.A significant upward shift in plant species optimum elevation during the 20th century[J].Science,2008,320:1768-1771.
猜你喜欢
水土保持研究(2022年1期)2022-12-27
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
青年文学家(2021年16期)2021-08-05
生态学报(2021年4期)2021-04-08
少儿美术(2020年8期)2020-11-06
林业调查规划(2020年3期)2020-06-03
中南林业科技大学学报(2018年10期)2018-07-27
现代园艺(2017年13期)2018-01-19
