
三倍体毛白杨地上凋落物对林龄的响应1)
2010-08-08 07:22:44宋曰钦翟明普贾黎明
东北林业大学学报 2010年3期
宋曰钦 翟明普 贾黎明
(安徽省黄山学院,黄山,245041) (教育部省部共建森林培育与保护重点实验室(北京林业大学))
纸浆林是 19世纪在欧美国家首先出现的一种工业原料林,经过多年的发展,目前已形成“高密度、短轮伐期、全树利用”的经营理念。这种经营理念对林地的养分管理提出了更高的要求。森林凋落物是指森林生态系统内、由生物组分产生并归还到林地表面,作为分解者的物质和能量来源以维持生态系统功能的所有有机质的总称[1]。森林凋落物是森林生态系统的重要组成部分,是森林生态系统中养分循环的重要一环,在恢复和增加土壤肥力、改善树木营养和提高森林生产力方面起到十分重要的作用,因此森林凋落物历来是森林生态学、生物地球化学和森林土壤学的重要研究内容。目前,在凋落物的总量、组成、动态变化,养分分解动态及凋落物能量的归还等方面已进行了大量的研究[2-6]。
三倍体毛白杨是朱之悌等采用细胞染色体部分替换和染色体加倍等技术,对毛白杨施加遗传改良获得的新品种[7],具有速生、优质、抗病等优点,尤其是优良的材性非常适合制浆造纸的需要,因而在我国林纸一体化工程建设中得到广泛栽植。目前对三倍体毛白杨的研究主要集中在育种技术及其形成机理[8]、材质及制浆造纸工艺[9]、造林技术及病虫害防治[10-11]等方面,有关养分循环及能量流动方面的研究尚未见报道。
1 试验地概况
试验地点位于山东高唐县,地理位置为东经116°01′~116°28′,北纬 36°39′~ 37°01′,地处黄河下游,属黄河冲积平原。高唐县属暖温带半干旱季风大陆性气候,具有明显的季节变化和季风气候特性。春旱多风,夏热多雨,秋爽易旱,冬寒少雪。年降水量 306.0~975.9mm,年平均气温 13.1℃,历年极端最高气温为41.2℃,极端最低气温-20.8℃。气候资源中热量,光照充足,水资源欠缺。年日照总时数 4 433.5h,全年无霜期204 d。
2 材料与方法
根据造林设计的档案记录,选取 4个不同林龄的三倍体毛白杨纸浆林作为研究对象。这4个林分均采用1年生根萌苗于春季造林,带状配置,带间距离为 8 m,两行为 1带,行间距离为 2m,株距为 1m。整地方式为机械穴状,穴直径为 1.2 m,深 1m,造林前以复合肥为基肥施于穴底部,回填少量表土后栽植苗木。其余情况见表1。
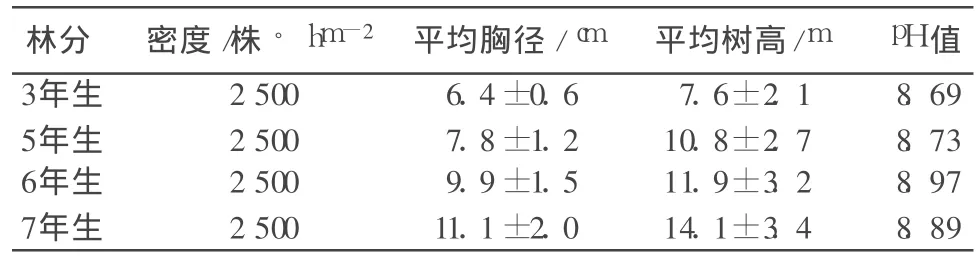
表1 试验林分基本情况
凋落物的收集方法:于 2008年 3月中旬在每一林分内确定 3个固定标准地,每块样地面积为 400m2,要求样地内的林木不少于100株。在每一样地布置 6个落叶收集器,其中 2个位于行间,2个位于带间,2个位于中间。收集器由 80目的尼龙网制成,呈圆锥型,高 70 cm,底圆直径 1.12m,以 4根支柱固定,尼龙袋离地面 10 cm。每月中旬收集样品,在室内除去鸟粪、昆虫尸体等杂物以后进行分类,分为叶、枝、芽及其他(花序、鳞片)成分,分别于 85℃条件下烘 48h,至恒质量,称质量,计算每月收集到的不同凋落物各组成部分的量及总量,并保留部分样品进行N、P营养元素及热值的测定。
分析方法:将每月保留的叶、枝及其他成分粉碎后过筛,保留在广口瓶中。采用凯氏定氮法测定氮;采用钼锑抗比色法测定磷,具体操作方法按国家标准LY-T 1269-1999及LY-T 1270-1999进行;热值采用 PARR 3600氧弹热量仪测定。
3 结果与分析
3.1 不同林龄三倍体毛白杨凋落物总量的变化
从 2008年 4月至 2009年 3月之间收集的不同林龄的三倍体毛白杨凋落物总量见表2。从表2可以看出,随着林龄的增加,地上凋落物的总量也增加,3年生林分共收集 216.03±59.73g◦m-2◦a-1,而 7年生林分凋落物总量达到 482.38±101.34g◦m-2◦a-1,7年生林分的年凋落物总量是 3年生林分的 223.39%,增加了 1倍多。方差分析结果表明,不同林龄林分的凋落物总量之间存在显著性差异(P=0.024 132),除6年生林分与 7年生林分之间差异不显著以外,其余林龄林分的凋落物总量的差异均达到显著(P<0.05)水平,这是由于 3~7年生三倍体毛白杨处于速生期,为维持个体的快速生长,需要不断增加叶面积以合成更多的同化产物,因此叶量也持续增加,从而使得凋落物的数量增加。这种凋落物总量随林龄增加而显著增加的现象,在其他研究中也有报道,李庆云等也发现在 5、7、9、11年生这 4个林龄阶段,杨树的凋落物总量分别为 0.97、2.14、4.82、6 t/hm-2◦ a-1,表现为随林龄增加而增加的趋势[12]。
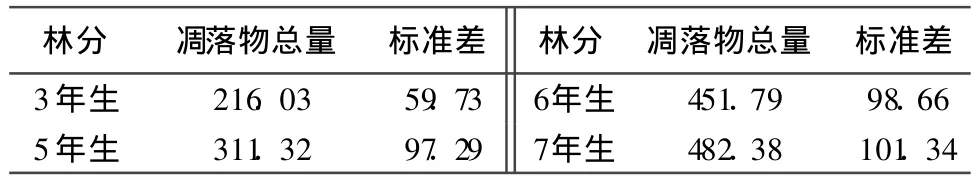
表2 不同林龄三倍体毛白杨 1 a中凋落物总量 g◦m-2◦a-1
3.2 不同林龄三倍体毛白杨凋落物的组成差异
森林以凋落物形式归还林地养分需要经过分解才能实现,尽管影响分解的因素很多,但凋落物的组成及性质是决定因素,因而研究不同林龄凋落物的组成可以预知凋落物的分解速度。从收集的凋落物总体来看,三倍体毛白杨的凋落物中以落叶为主,落叶所占的比例随林龄的增大而降低(表3),其中 3年生林分落叶所占比例为 98.64%,到 7年生林分落叶所占比例为95.75%;枝的比例随林龄增加而增加,3年生林分的凋落物中只有 1.2%为枝,到 7年生林分时这一比例上升到 4.06%;芽、鳞片及花絮等其他部分的产生时间短、量少,因此所占比例很低。

表3 不同林龄三倍体毛白杨凋落物组成 g◦m-2◦a-1
3.3 不同林龄三倍体毛白杨凋落物的月动态变化
植物的凋落具有周期性的变化规律[13]。从不同林龄三倍体毛白杨凋落物总量的月变化趋势(表4)来看,呈一条明显的单峰曲线,2008年 4—7月份,形成的凋落物量小,各月份之间变化幅度小,这种趋势一直持续到 8月份,只有 5年生林分出现例外,8月份的凋落物量明显上升,在 9月份形成高峰,其余林分在 9月份开始上升后,于 10月份达到峰值。通过鉴定,造成 5年生林分凋落物峰值出现时间提前的原因是由于叶片感染病菌造成。从不同组分的出现时间来看,2008年 5月份就可以收集到落叶,一直持续到 2008年的 11月份。不同林龄枯枝出现的时间不同,其中 7年生林分在 2008年4— 5月份、7— 11月份,以及 2009年 1— 3月份均可收集到枯枝,而 2008年的 6月份、12月份却未收集到;6年生林分在4— 5月份及 8— 9月份,2009年的 2月份、3月份可收集到枯枝,其余月份均未收集到枯枝;5年生林分则是在 2008年 4—8月份及 2009年的 2月份可收集到枯枝;3年生林分仅在2008年 4、5、12月份及 2009年 2月份可收集到枯枝。造成这种差异的原因可能与不同林分枯枝形成的状况、当地风速等因素有关。芽的出现时间集中在展叶后的 5—6月份;5月份从 5、6、7年生林分中可收集到花絮。
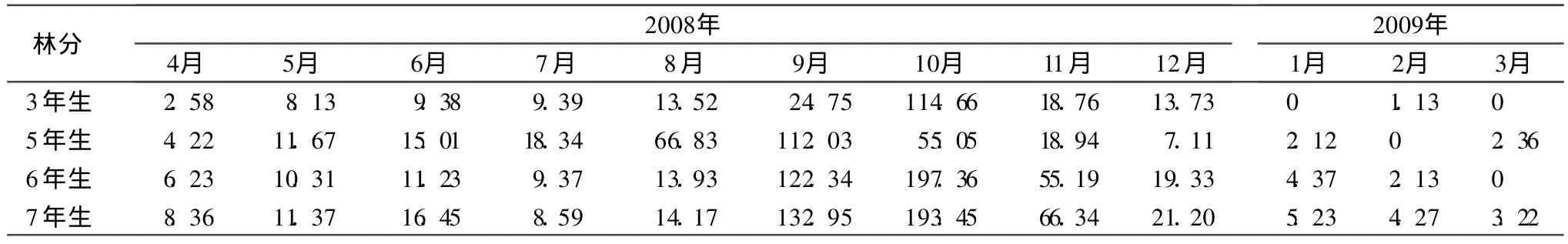
表4 不同林龄三倍体毛白杨 1a中不同月份凋落物量 g◦m-2◦a-1
3.4 不同林龄三倍体毛白杨凋落物 N、P归还动态
森林凋落物在整个森林生态系统中最重要的功能之一是向林地归还养分,归还的具体数量受凋落物总量及凋落物中不同养分量的影响。从不同林龄的三倍体通过落叶可能归还的 N、P量来看(表5),除 7年生林分归还的 N低于 6年生林分外,其余均表现为随林龄增加而增加的趋势,3年生林分通过落叶归还林地的 N为 1.82g◦m-2◦a-1,归还的 P为 0.57g◦m-2◦a-1;5年生林分的落叶能归还林地的N为 2.14g◦m-2◦a-1,P为 0.79g◦m-2◦a-1;6年生林分的落叶归还林地的 N为 2.87g◦ m-2◦ a-1,P为 1.08g◦ m-2◦ a-1;7年生林分的落叶归还林地的 N为 2.80g◦m-2◦a-1,归还的 P为 1.09g◦m-2◦a-1。从不同月份的动态变化来看,呈单峰曲线,与落叶量的月变化相同,这是由于虽然不同月份的落叶中养分的量存在一定的差异,但这种差异小于落叶量的差异,因此落叶量越大,归还的养分越多。
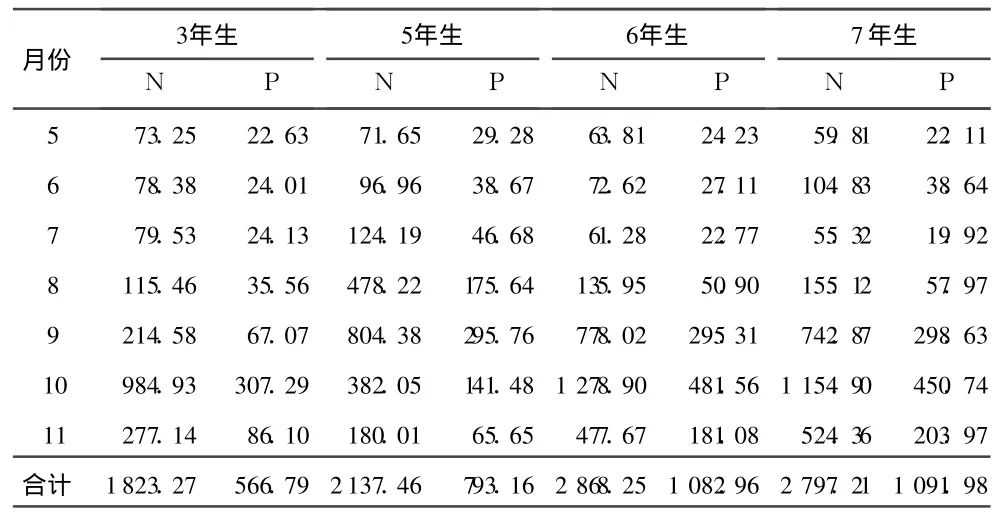
表5 不同林龄三倍体毛白杨枯落叶归还林地 N、P的总量mg◦ m-2◦ a-1
由不同月份凋落枝条的量及其 N、P的量,可以得到不同林龄三倍体毛白杨纸浆林凋落枝归还林地的 N和P的量(表6)。

表6 不同林龄三倍体毛白杨凋落枝归还林地 N、P总量mg◦ m-2◦ a-1
由表5及表6中相应林龄的三倍体毛白杨通过落叶和落枝归还的N、P量相加,则不同林龄的三倍体毛白杨每年通过地上凋落物归还的 N、P分别为:3年生林分归还 N 1 838.39 mg◦ m-2◦ a-1,归还 P 580.26mg◦ m-2◦ a-1,其中落叶归还的 N占 99.18%,归还的 P占 97.68%;5年生林分归还 N 2163.42、P 815.95mg◦ m-2◦ a-1,其中落叶归还的 N占 98.80%,归还的 P占 97.21%;6年生林分归还 N 2 922.99、P 1131.44 mg◦m-2◦a-1,其中落叶归还的 N占 98.13%,归还的 P占 95.72%;7年生林分归还 N 2888.29、P 1 171.65mg◦ m-2◦ a-1,其中落叶归还的N占 96.85%,归还的P占 93.20%。
3.5 不同林龄三倍体毛白杨凋落物能量归还的动态变化
凋落物在为林地提供养分的同时,也提供了分解者(土壤动物、微生物)活动所需的能量,从而直接影响到凋落物的分解和养分的释放,因此研究凋落物能量归还的动态可以深入认识森林中能量流动的状况,也可以更深入认识分解者在凋落物分解中的作用[4]。从不同林龄的三倍体毛白杨归还林地的能量变化来看(表7),基本与凋落物质量的月动态相似,表现为单峰曲线,除 5年生林分的峰值出现在 9月以外,其他林分均在10月份。这是由于虽然不同凋落物的热值不同,但 3~7年生林分凋落物组成以落叶为主,枝、芽及花絮等在凋落物的总量中所占比例低,而凋落物的单位热值在不同月份间差异较小,其热值的差异不足以改变整个趋势。从单位质量的热值变化来看,同一林龄不同器官的热值变化顺序为花序>叶>枝,这与以往报道一致[14],不同林龄相同器官之间存在一定差异,但没有发现规律,如7年生树叶热值为1.81×104J/g,6年生树叶热值为 1.83×104J/g,5年生树叶热值为 1.74×104J/g,3年生树叶热值为 1.84×104J/g。从归还的总量来看,不同林龄的林分归还总量随林龄增大而增加,其中 3年生林分归还林地的能量为 3.96×106J◦m-2◦a-1,5年生林分则为 5.43×106J◦m-2◦a-1,6年生林分为 8.20×106J◦m-2◦a-1,7年生林分达到 8.89×106J◦ m-2◦ a-1。
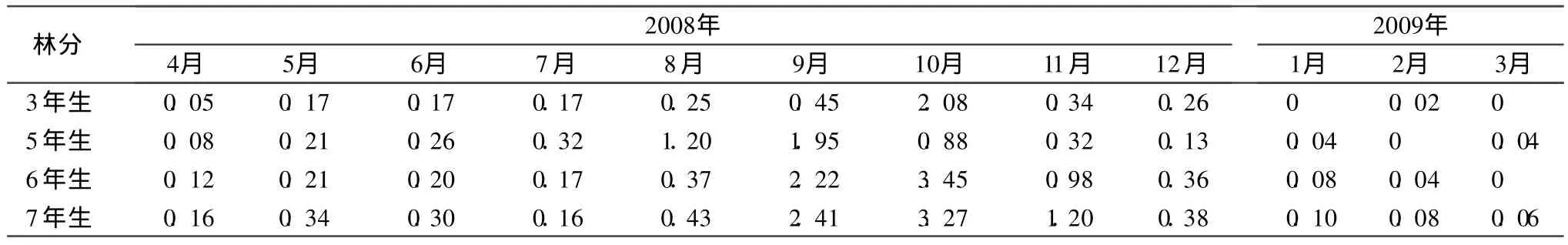
表7 不同林龄三倍体毛白杨凋落物归还能量的月变化 106 J◦m-2◦a-1
4 结论与讨论
一般认为凋落物的总量是由自身生物学特性决定的,不同树种凋落物数量不同[15],其他因素也影响到凋落物的量,如在不同林分密度下,凋落物量及归还养分量都不相同,表现为随着林分密度增加而增加[16]。还有一些研究指出,全年落叶量与林分的树干基部断面积呈正相关[17],另外,经营措施如间伐等也对凋落物总量产生影响[18]。本研究中4个不同林龄(3~7年生)的三倍体毛白杨年归还总量为(216.03±59.73)~(482.38±101.34)g◦ m-2◦a-1,归还凋落物总量随林龄的增加而增加,由于这 4个不同林龄的林分立地条件相似,林分密度一致,经营措施基本相同,因此凋落物总量的差异主要是由林龄的差异造成的。
从凋落物组分来看,在 4个不同林龄的林分中均以叶为主,这与以往报道的结果一致,但落叶所占比例为 95.75%~98.64%,高于文献中报道的比例[19],这可能与本研究中 4个林分的最大林龄仅为 7a,而且三倍体毛白杨自身细枝数量少的分枝特性也影响到枯枝形成量,因此所占比例较小。
在 1a中不同林龄的林分凋落物的月动态变化均为单峰曲线,除 5年生林分因叶片感染病菌于 9月份形成凋落高峰以外,其余林分均在 10月份形成凋落高峰。这种由于病虫害等突发因素对凋落节律的影响以往也有报道[19]。
不同林龄的林分凋落物归还 N、P的总量随林龄增加而增加,其中归还的 N为 1.84~2.89g◦m-2◦a-1,归还的 P为0.58~1.17g◦m-2◦a-1,归还的 N、P的月动态变化规律与凋落物量的动态一致。从输入林地的总能量来看,在 1a中不同林龄三倍体毛白杨地上凋落物归还林地的能量总量随林龄的增加而增加,3年生林分归还的能量为 3.96×106J◦m-2◦a-1,5年生林分归还的能量为 5.43×106J◦m-2◦a-1,6年生林分归还的能量为 8.20×106J◦m-2◦a-1,到 7年生林分归还的能量达到 8.89×106J◦m-2◦a-1。 7年生林分通过凋落物归还林地的能量是 3年生林分的 224.49%。与其他生态系统的能量流相比,即使林龄为 7 a的三倍体毛白杨林分通过凋落物归还的能量也小于福建亚热带雨林[14](16.811×106J◦m-2◦a-1)、福建杉木林 (10.84 ×106J◦ m-2◦ a-1)和杉木观光混交林[20](11.24×106J◦m-2◦a-1),但高于武夷山的甜储林群落[21](7.679×106J◦m-2◦a-1),造成不同生态系统类型之间凋落物总能量差异可能与不同生态系统类型的凋落物数量、组成和热值有关。
[1] Norris M D,Blair JM,Johson LC.Land cover change in eastern Kansas:litter dynamics of closed-canopy eastern redcedar forests in tall grass prairie[J].Canadian Journal of Botany,2001,79(2):214-222.
[2] Bernal S,Butturini A,Nin E,et al.Leaf litter dynamics and nitrous oxide em ission in a Mediterranean riparian forest:implications for soil nitrogen dynamics[J].Journal Environmental Quality,2003(32):191-197.
[3] Isaac S R,Nair M A.Litter dynam ics of six multipurpose trees in a homegarden in south Kerala,India[J].Agroforestry Systems,2006(67):203-213.
[4] 何宗明,陈光水,王巧珍,等.33年生福建柏人工林群落能量的研究[J].应用与环境生物学报,2003,9(6):569-573.
[5] 张雪萍,黄初龙,李景科.赤子爱胜蚓对森林凋落物的分解效率[J].生态学报,2005,5(9):2027-2033.
[6] 黄钰辉,官丽莉,周国逸,等.西双版纳热带季雨林和哀牢山中山湿性常绿阔叶林优势植物及地表凋落物层的热值[J].植物生态学报,2007,31(3):457-463.
[7] 朱之悌,林惠斌,康向阳.毛白杨异源三倍体 B301等无性系选育的研究[J].林业科学,1995,31(6):499-505.
[8] 康向阳,张平东,高鹏,等.秋水仙碱诱导白杨三倍体新途径的发现[J].北京林业大学学报,2004,26(1):5-8.
[9] 蒲俊文,宋君龙,姚春丽.三倍体毛白杨纤维形态变异的研究[J].北京林业大学学报,2002,24(2):68-72.
[10] 薛睿,贺斌,薛文瑞.甘肃干旱荒漠区纸浆林三倍体毛白杨营建试验[J].陕西林业科技,2006(2):12-16,36.
[11] 林仲桂,张仁福,钟福生,等.危害三倍体毛白杨的害虫种类、发生与防治研究[J].湖南环境生物职业技术学院学报,2006,12(1):1-4.
[12] 李庆云,万猛,樊巍,等.黄淮海平原农区杨树人工林生物量和生产力研究[J].河南科学,2008,26(2):434-437.
[13] 张翼,汪有科,吴钦孝.黄土高原几种主要森林类型的凋落及其过程的比较研究[J].水土保持学报,2001,15(5):91-94.
[14] 林鹏,邵成.福建和溪亚热带雨林凋落物能流的研究[J].厦门大学学报:自然科学版,1995,34(6):987-991.
[15] 俞益武,吴家森,姜培坤,等.湖州市不同森林植被枯落物营养元素分析[J].浙江林学院学报,2002,19(2):153-156.
[16] 姚瑞玲,丁贵杰,王胤.不同密度马尾松人工林凋落物及养分归还量的年变化特征[J].南京林业大学学报:自然科学版,2006,30(5):83-86.
[17] Berg B,Meentemeyer V.Litter fall in some European coniferous forests as dependent on climate:a synthesis[J].Canadian Journal of Forest Research,2001(31):292-301.
[18] Inagaki Y,Sakai A,Kuramoto S,et al.Inter-annual variations of leaf-fall phenology and leaf-litter nitrogen concentration in a hinoki cypress(Chamaecyparis obtusa Endlicher)stand[J].Ecological Research,2008,23(6):965-972.
[19] 翟明普.北京西山地区油松元宝枫混交林生物量和营养元素循环的研究[J].北京林学院学报,1982(4):67-79.
[20] 杨玉盛,陈光水,林瑞余,等.杉木观光木混交林群落的能量生态[J].应用与环境生物学报,2001,7(6):536-542.
[21] 林益明,林鹏,李振基,等.福建武夷山甜储群落能量的研究[J].植物学报,1996,38(12):989-994.
猜你喜欢
中国水产(2024年3期)2024-04-11 07:36:00
科学养鱼(2021年11期)2022-01-11 11:21:10
防护林科技(2020年10期)2020-12-22 11:46:42
新农民(2020年21期)2020-12-08 13:59:06
食品与生活(2019年12期)2019-05-15 15:01:41
生命科学研究(2018年1期)2018-05-29 01:12:56
东坡赤壁诗词(2018年1期)2018-03-31 09:10:10
河南林业科技(2017年1期)2017-06-10 08:00:39
广东农业科学(2016年7期)2016-10-08 05:56:40
防护林科技(2016年9期)2016-09-27 01:34:21

- 东北林业大学学报的其它文章
- 城市森林的近自然林经营技术方案