
不同温度和浓度下核型多角体病毒对甜菜夜蛾的致病力
2010-06-19 07:32:42徐彩霞韩日畴
长江蔬菜 2010年18期
徐彩霞,韩日畴
(广东省昆虫研究所,广州,510260)
甜菜夜蛾是一种抗药性发展快的害虫,已对多类化学农药产生高度抗性,因此生物防治方法的研究不断普遍和深入。甜菜夜蛾生物防治方法的研究主要集中在病原微生物上。Chambers对Bacillus thuringiensis中的一种新型杀虫晶体蛋白的基因进行了相关研究[3]。Moulton又报道了一种从Bt分离出来的结晶蛋白的杀虫机理[2]。核型多角体病毒是调节甜菜夜蛾自然种群的重要因子。自1949年Steinhaus首先在美国加利福尼亚州分离到一株甜菜夜蛾核型多角体病毒(SeNPV)以来[5],国内外开展了很多分离鉴定[6]、毒力测定[7,8]、致病机理[9]、病毒制剂及田间应用等方面的研究[10]。SeNPV在大田的使用效果受到浓度、温度等多种因素的影响[11],防治效果不尽相同。本文研究了不同温度和浓度下SeNPV对甜菜夜蛾致病力的影响,以期为田间应用提供参考。
1 材料与方法
1.1 供试病毒
甜菜夜蛾核型多角体病毒来自实验室保存。在室内用甜菜夜蛾幼虫增殖一次后,将病死虫尸匀浆,双层纱布过滤,差速离心3次后,取少量病毒悬浮液,在显微镜下用血球计数板计数,计算病毒含量后于4℃冰箱中保存。
1.2 供试昆虫
甜菜夜蛾3龄幼虫,采自田间,经多代人工饲料室内饲养后供试验。
1.3 试验方法
①不同浓度SeNPV对甜菜夜蛾致病力的影响用PBS缓冲液将SeNPV悬浮液配成1×107PIBs/mL、1×106PIBs/mL、1×105PIBs/mL、1×104PIBs/mL、1×103PIBs/mL 共5种浓度。番薯叶(约3 cm2)洗净后,在每片叶上滴入5 μL不同浓度的病毒溶液,自然晾干后置于灭过菌的培养皿(直径9 cm)中,加入1头3龄幼虫,24 h后叶片被全部吃完,然后用无毒的番薯叶饲喂,以10头幼虫为1个重复,每处理重复3次。以饲喂无毒的番薯叶为对照。所有培养皿统一放置在空调养虫室的养虫笼内,温度(25±1)℃,相对湿度(65±5)%。 每 24 h 观察记录幼虫的病死数和病死情况,并更换新鲜的番薯叶供幼虫食用,共观察10 d(或至幼虫全部死亡)。
②不同温度下SeNPV对甜菜夜蛾的致病力 用PBS缓冲液将SeNPV病毒配制成浓度为1×106PIBs/mL的悬浮液。洁净的番薯叶剪成面积约3 cm2,在每片叶上滴入5 μL病毒悬浮液,自然晾干后放于灭过菌的培养皿(9 cm)内,加入1头3龄幼虫,24 h后叶片被全部吃完,然后用无毒的番薯叶饲喂,以10头幼虫为1个重复,每处理重复3次。以饲喂无毒的番薯叶为对照。培养皿放置在人工气候箱内,温度设置为15,20,25,30,35±1℃,相对湿度(65±5)%。 温度为(15±1)℃时,幼虫取食量较小,因而试验用较小面积(约2 cm2)的番薯叶。每24 h观察记录幼虫的发病和病死情况,并更换新鲜的番薯叶供幼虫食用,共观察10 d(或至幼虫全部死亡)。
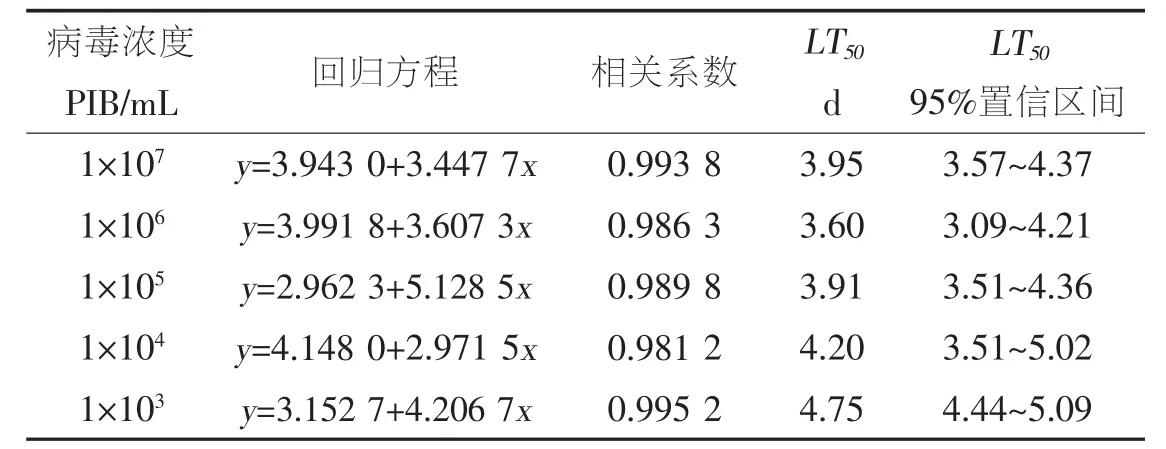
表1 不同浓度SeNPV对甜菜夜蛾3龄幼虫的毒力回归方程和LT50
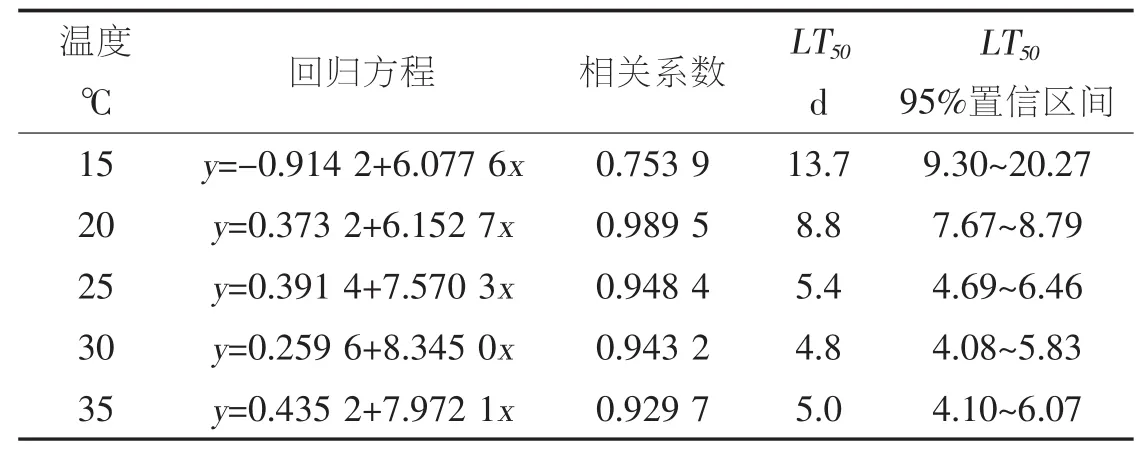
表2 不同温度下SeNPV对甜菜夜蛾3龄幼虫的毒力回归方程和LT50
1.4 数据分析
按照以下公式计算不同病毒浓度和温度下幼虫的累计死亡率。
累计死亡率 (%)=(在一定时间内总死亡数/初始虫数)×100%;累计校正死亡率 (%)=(处理累计死亡率-对照累计死亡率)/(1-对照累计死亡率)×100%。
以时间的对数值为X,死亡率概率值为Y,计算不同浓度和温度下的毒力回归方程和致死中时间(LT50)。用单因素方差分析比较不同浓度或温度下不同时间幼虫死亡率的差异 (SPSS 13.0,Turkey's test),分析不同环境因子对甜菜夜蛾核多角体病毒致病力的影响。
2 结果与分析
2.1 不同浓度下核型多角体病毒对甜菜夜蛾致病力的影响
图1显示了不同浓度下,SeNPV病毒对甜菜夜蛾幼虫的累计死亡率。由图1可以看出,随着处理时间的延长,幼虫死亡率增加,感染10 d后,浓度为1×107PIBs/mL、1×106PIBs/mL、1×105PIBs/mL、1×104PIBs/mL、1×103PIBs/mL时,累计死亡率分别为83.3%,83.3%,86.7%,90.3%,96.0%,差异不显著(F=0.917,df=4,10,p=0.491)。
根据图1的结果计算不同浓度下SeNPV对甜菜夜蛾3龄幼虫的毒力回归方程和LT50见表1。由表1可知,感染时间受病毒浓度的影响不大,病毒浓度为 1×106PIBs/mL 时,LT50最小,为 3.60 d,相差仅1.15 d。
2.2 不同温度下核型多角体病毒对甜菜夜蛾的致病力的影响
图2显示了不同温度下甜菜夜蛾3龄幼虫感染病毒不同时间后的累计死亡率。温度显著影响甜菜夜蛾病毒的感染力。感染10 d后,温度为15,20,25,30,35℃时, 甜菜夜蛾幼虫的累计死亡率分别为 53.3%,66.7%,91.4%,96.7%,96.7%,差异显著(F=9.075,df=4,10,p=0.002)。
温度显著影响SeNPV对甜菜夜蛾的致病力,温度为 15,20,25,30,35℃时, 病毒对 3 龄幼虫的LT50分别为 13.7 d,8.8 d,5.4 d,4.8 d,5.0 d。 随着温度的升高,LT50逐渐减小。温度为15℃时LT50最小,温度为30℃时,LT50最大,约是前者的3倍。温度为35℃时,LT50较温度30℃时略微增大,说明温度过高可能对病毒致病能力存在一定的抑制作用。
3 结论与讨论
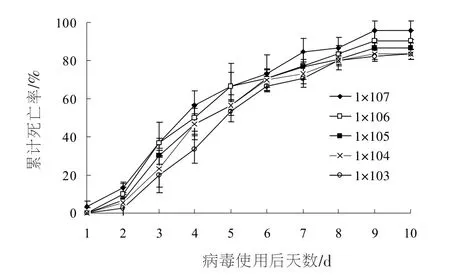
图1 不同浓度下甜菜夜蛾3龄幼虫感染SeNPV的累计死亡率
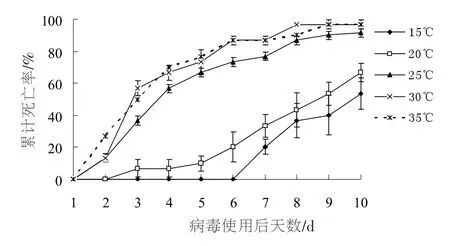
图2 不同温度下甜菜夜蛾3龄幼虫感染SeNPV的累计死亡率
①病毒浓度对SeNPV致病力的影响不明显,病毒浓度为 1×107PIBs/mL、1×106PIBs/mL、1×105PIBs/mL、1×104PIBs/mL、1×103PIBs/mL 时,感染 10 d 后的累计死亡率均达到80%以上,结果差异不显著,这与前人的研究结果不一致。前人指出,在一定范围内,不同浓度SeNPV感染甜菜夜蛾2龄幼虫后,幼虫死亡率随着浓度的增大而升高[12],这可能是因为本试验所用的幼虫为生物公司人工饲养的昆虫,对病毒更为敏感。
②环境温度显著影响SeNPV对甜菜夜蛾3龄幼虫的致病力,随着温度的升高,病毒对目标昆虫的毒力明显增强。温度低于25℃时,病毒对幼虫的致病力明显减弱,30℃是幼虫感染病毒的最适温度,温度升高至35℃时,病毒毒力反而略微降低。蒋杰贤等报道29℃是病毒的最适侵染温度,本文的研究结果与其一致[13]。温度为20℃时的校正死亡率为72.22%,本文结果与其相当;温度为35℃时死亡率仅为53.13%[13],本文结果与其出入较大,这可能与虫源、病毒株及试验方法有关。
[1]Mascarenhas V J,Graves J B,Leonard B R,et al.Susceptibility of field populations of beet armyworm(Lepidoptera:Noctuidae)to commercial and experimental insecticide[J].Journal of Economic Entomology,1998,91,827-833.
[2]Moulton J K,Pepper D A,Jansson R K,et al.Pro-active management of beet armyworm(Lepidoptera:Noctuidae)resistance to tebufenozide and methoxyfenozide:Baseline monitoring,risk assessment,and isolation of resistance[J].Journal of Economic Entomology,2002,95:414-424.
[3]Chambers J A,Jehen A,Gilbert M P,et al.Isolation and characterization of a novel insecticidal crystal protein gene fromBacillus thuringiensissubsp.aizawai [J].Journal of Bacteriology,1991,173:3 966-3 976.
[4]Kalman S,Kiehne K L,Libs J L,et al.Cloning of a novel CryIC-type gene from a strain ofBacillus thuringiensis subsp.gallerae[J].Applied and Environmental Microbiology,1993,59:1 131-1 137.
[5]Steinhaus E A.Nomenclature and classification of insect viruses[J].Bacteriological Reviews,1949,13:203-223.
[6]Hara K,Funakoshi M,Kawarabata T.In vivo and in vitro characterization of several isolates of Spodoptera exigua nuclear polyhedrosis virus [J].Acta Virologica,1995,39:215-222.
[7]Gelernter W D,Toscano N C,Kido K,et al.Comparison of a nuclear polyhedrosis virus and chemical insecticides for control of the beet armyworm (Lepidoptera:Noctuidae)on head lettuce[J].Journal of Economic Entomology,1986,79:714-717.
[8]张海元,梅春蕾,张忠信.SeMNPV两个分离株的生物活性比较及病毒杀虫剂的应用[J].中国病毒学,2005,20(6):656-659.
[9]Wilson J A,Hill J E,Kuzio J,et al.Characterization of the baculovirusChoristoneura fumiferanamulticapsid nuclear polyhedrosis virus p10 gene indicates that the polypeptide contains a coiled-coil domain [J].Society for General Microbiology,1995,76:2 923-2 932.
[10]Smits P H,van de Vrie M,Vlak J M.Nuclear polyhedrosis virus for control ofSpodoptera exigualarvae on glasshouse crops [J].Entomologia Experimentlis et Applicata,1987,43:73-80.
[11]郑常格,李广宏,陆小军,等.环境因子对SeNPV杀虫剂防治效果的影响[J].昆虫天敌,2006,28(2):66-70.
[12]刘琴,徐健,殷向东,等.甜菜夜蛾核型多角体病毒对甜菜夜蛾幼虫的毒力测定[J].江苏农业科学,2006,1:60-61.
[13]蒋杰贤,王冬生,曾爱平,等.温度对甜菜夜蛾核型多角体病毒流行的影响 [J].生态学报,2004,24 (8):1 726-1 730.
猜你喜欢
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:04
中老年保健(2021年11期)2021-08-22 03:16:00
基层中医药(2021年11期)2021-06-05 06:54:16
小哥白尼(趣味科学)(2021年12期)2021-03-16 05:40:44
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
作文周刊·小学二年级版(2016年17期)2017-05-27 02:36:52
中国糖料(2016年1期)2016-12-01 06:49:04
小学生导刊(2016年34期)2016-08-22 12:30:38
哈尔滨医药(2015年2期)2015-12-01 03:57:21
中国当代医药(2015年9期)2015-03-01 02:02:12
