
RHDV CD株 VP60主要抗原表位基因的克隆与序列分析
2010-05-18 11:14杨金生
中国动物检疫 2010年8期
隋 慧,杨金生
(1.辽宁医学院畜牧兽医学院,辽宁 锦州 121001;2.吉林省兽医科学研究所,吉林 长春 130062)
兔出血症(Rabbit haemorrhagic disease,RHD)是1984年由我国首先发现的家兔的一种烈性毁灭性病毒性传染病,病原为RHDV。RHDV属于杯状病毒科兔病毒属。RHDV基因组为单股正链RNA,由7437个核苷酸组成。RHDV基因组含有2个ORFs。除了基因组RNA外,RHDV还包含一个2.2 kb的亚基因组。ORF1编码一个多聚蛋白。该多聚蛋白被病毒蛋白酶进一步分解为衣壳蛋白(65 KD)和多个非结构蛋白,其中衣壳蛋白为病毒的主要结构蛋白,称VP60。体外翻译表明亚基因组也编码VP60。VP60基因是RHDV的唯一结构蛋白基因,在病毒的诱导机体免疫反应中起主要作用,其核苷酸的极端保守性正是目前RHDV仅有一个血清型的原因所在,也是现有RHDV疫苗能提供较好保护能力的重要原因之一,同时也为科研人员研制基因工程疫苗带来了极大的便利。VP60蛋白从N端到 C 端可分 A、B、C、D、E、F 6 个区域,西班牙学者E.Viaplana等证实了RHDV的A、B段包括最初的175个氨基酸比VP60的其它部分具有更高的抗原性,这个区域在病毒的衣壳中具有高度保护性。
1 材料与方法
1.1 材料
RHDV CD株,由辽宁省益康生物制品厂提供;反转录酶AMV RT、Rnasin、dNTPs、Ex Taq、 溴化乙锭(EB)、MarkerDL2000 均 为 大 连Takara公司产品;琼脂糖(Agarose)为Oxoid公司产品;DNA回收试剂盒为杭州Vitagene公司产品;LS Trizol试剂为Invitrogen公司产品;DEPC水购自宝泰克生物科技公司。
1.2 方法
1.2.1 引物设计
参照GenBank发表的RHDV基因序列,设计了扩增VP60主要抗原表位基因的上下游引物P1、P2。为方便后续工作,在上下游引物两端分别加EcoRI、XhoI酶切位点,送宝泰克生物技术公司合成。
上游引物 P1:5'-ATGAATT CGAGGGCAAAGCCCG-3'
下游引物 P2:5'-ATGCTCG AGTTAGGGACGCAAGTCTGG-3'
斜体加黑部分分别为EcoRI、XhoI酶切位点,引物P1、P2使用前按照说明书将其用灭菌三馏水稀释成工作浓度20~30 pmol/μL。
1.2.2 病毒增殖
取RHDV种毒10×稀释后,皮下接种成年易感家兔。待家兔发病死亡后,取其肝脏捣碎,加10×无菌生理盐水匀浆,冻融3次,10000 r/min离心10 min,取上清-20℃保存。
1.2.3 病毒RNA提取及RT-PCR
按照Trizol试剂盒方法提取RNA。反转录设为25 μL体系,下游引物 2 μL、病毒 RNA 10 μL,置 70℃ 5 min,冰浴 5 min。加入 10 mmol/L dNTP 3 μL、5 ×AMV RT Buffer 5 μL、Rnasin 1 μL (40 U)、AMV 反转录酶 2 μL、DEPC水 2 μL,置42℃水浴90 min,冰水浴5 min后立即进行PCR反应。PCR反应:模板(反转录产物)3 μL,上游引物 P1(20~30 pmol/μL)5 μL,下游引物 P2 (20~30 pmol/μL)6 μL,为dNPT (2.5 mM each)4μL,10×Ex Taq Buffer 5 μL,灭菌三馏水26.5 μL,Ex Taq (5 U/μL)0.5 μL。置PTC100型PCR扩增仪进行扩增,反应条件为:94℃预变性4 min;94℃35 s,50℃ 35 s,72℃ 1 min,30个循环后,72℃延伸10 min。产物经1%琼脂糖凝胶电泳分析。将纯化的PCR产物送Takara公司进行序列测定。
1.2.4 各RHDV株间同源性和系统发生分析
采用DNASTAR软件,对14株不同地区的RHDV进行核酸及推导氨基酸同源性分析。
2 结果
2.1 RHDV CD株VP60主要抗原表位基因的PCR扩增
RHDV接种易感兔后,家兔发病死亡,从兔肝组织匀浆中提取病毒RNA,采用RT-PCR方法扩增VP60主要抗原表位基因,产物经1%琼脂糖凝胶电泳分析,在525 bp处出现一条DNA带(图1)。
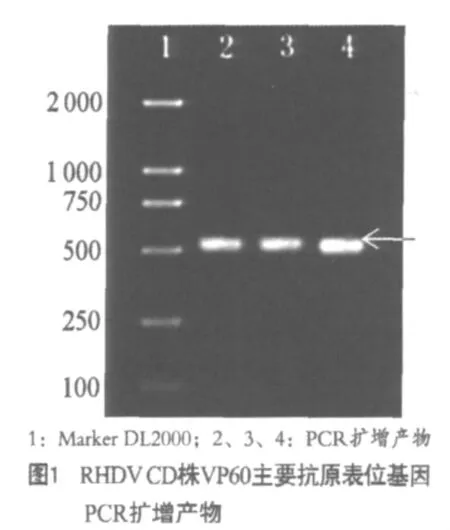
2.2 测序结果
测序结果显示,VP60主要抗原表位基因长525 bp(图2)。
2.3 核酸序列分析
2.3.1 CD株VP60主要抗原表位基因的核酸序列分析
RHDV CD株VP60主要抗原表位基因全长 525 bp,A、T、G、C 含量分别为 23.62%、22.10%、26.67%、27.62%,GC含量为54.29%。

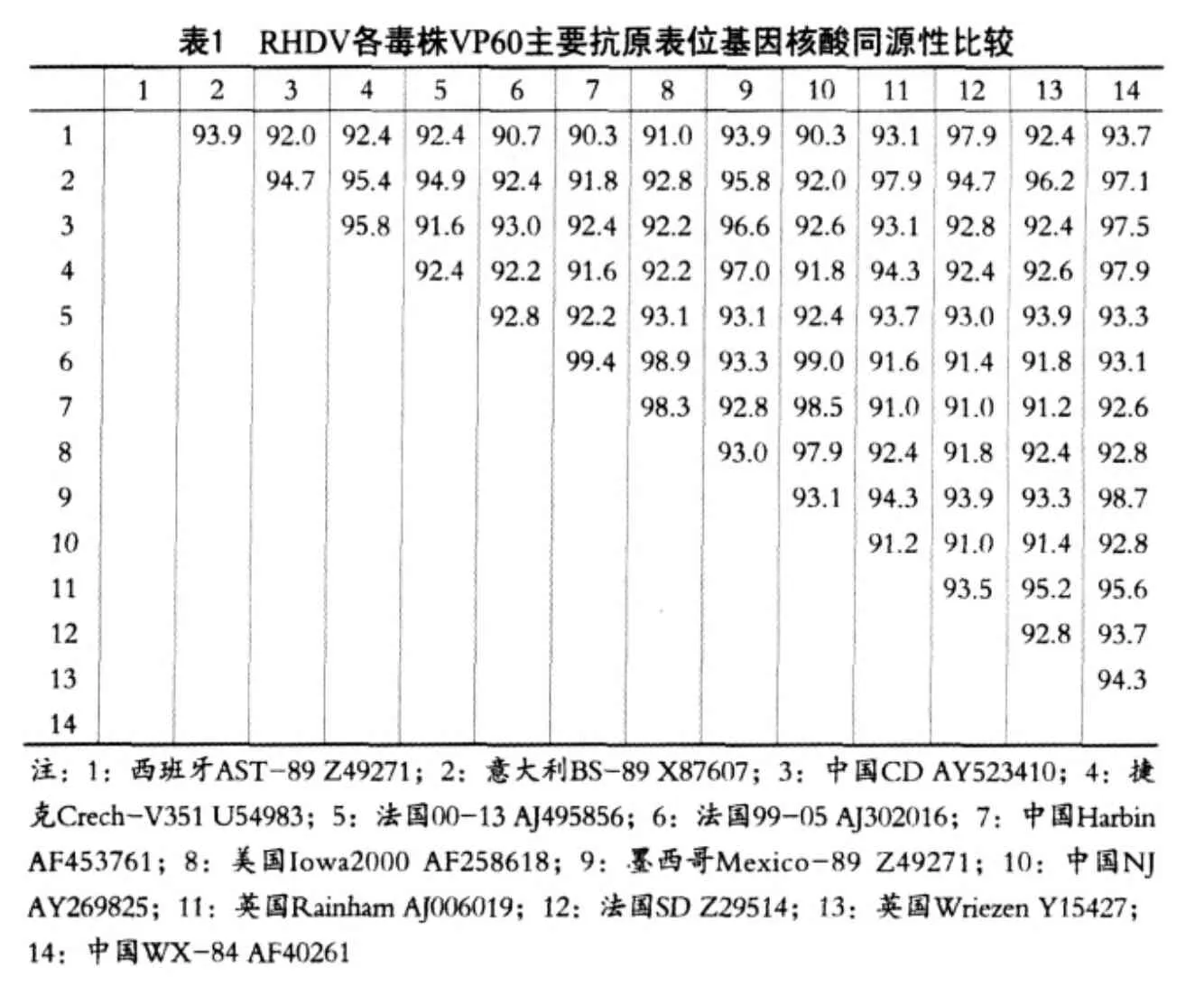
2.3.2 毒株之间核酸的同源性比较
RHDV CD株VP60主要抗原表位基因与其它13株来自中国(WX-84 AF402614、NJ AY269825、Harbin AF453761)株、法国(99-05 AJ302016、00-13 AJ495856、SD Z29514)株、墨西哥(Mexico-89 AF295785)株、意大利(BS-89 X87607)株、英国(Rainham AJ006019、Wriezen Y15427)株、西班牙 (AST-89Z49271) 株、捷克(Crech-V351U54983) 株 、 美 国(Iowa2000 AF258618) 株的 RHDV VP60同一序列进行了核苷酸序列同源性比较。结果发现,CD株与WX-84株同源性最高为97.5%,与其它毒株的同源性在92.0%~96.6%,其它毒株间的同源性在90.3%—99.4%(表1)。
?
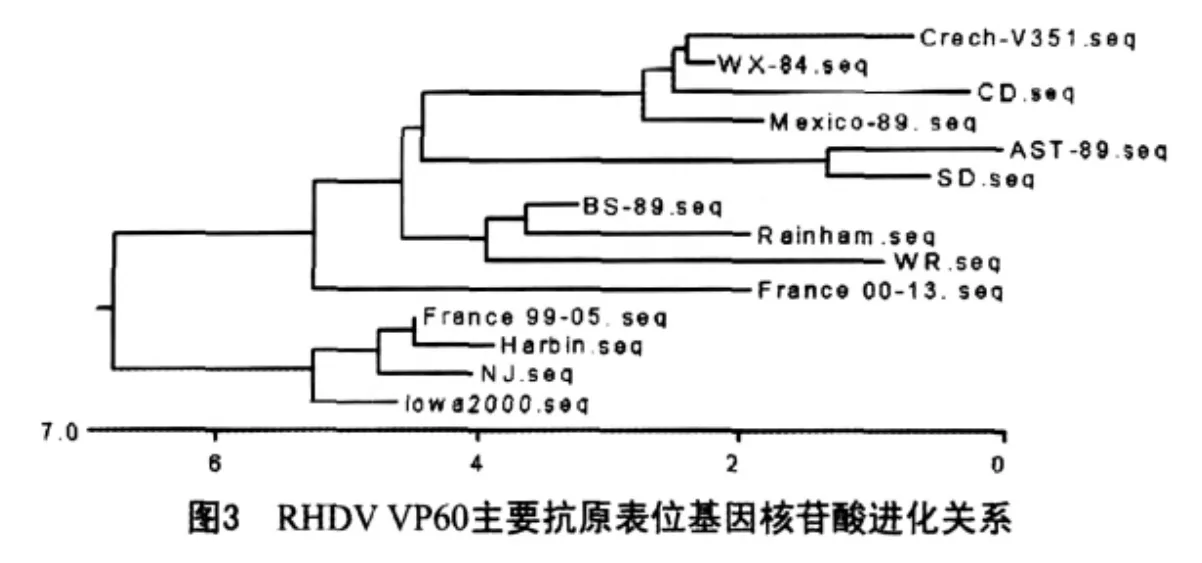
2.3.3 RHDV各毒株间核苷酸进化关系
如图3所示。
2.4 毒株之间氨基酸序列比较
2.4.1 毒株之间氨基酸同源性比较
RHDV CD株VP60主要抗原表位基因推导的氨基酸序列与其它13株来自中国(WX-84AF402614、NJ AY269825、Harbin AF453761) 株、法国(99-05 AJ302016、00-13 AJ495856、SD Z29514)株、墨西哥(Mexico-89 AF295785)株、意大利(BS-89 X87607)株、英国(Rainham AJ006019、Wriezen Y15427)株、西班牙(AST-89 Z49271)株、捷克(Crech-V351 U54983)株、美国(Iowa2000 AF258618)株的 RHDV VP60同一序列推导的氨基酸序列进行了同源性比较。结果发现,CD株与WX-84株、墨西哥Mexico-89株同源性最高,为99.4%;与其它毒株的同源性在96.6%~98.3%之间,其它毒株间的同源性在96.0%~100.0%之间(表2)。
2.4.2 RHDV各毒株间氨基酸进化关系
如图4所示。
3 讨论
?
RHD是二十世纪末新发现的一种对兔危害极大的传染病,一开始就引起人们极广泛的兴趣。由于RHDV不能在鸡胚上增殖,也难于在各种原代或传代细胞中稳定增殖,故至今尚未真正建立体外组织细胞的培养方法,所以目前广泛使用的疫苗为组织灭活苗,即取人工感染的病兔肝脏制成匀浆,加入甲醛等灭活剂进行灭活后使用。一般采取皮下或肌肉注射接种,具有良好的免疫效果。但目前随着家兔饲养成本的提高以及非免疫兔的减少,可用于制苗的感染材料日趋减少,使组织灭活苗的成本不断增加,加之组织灭活苗本身所不可避免的种种缺点以及动物福利等[1]问题,使得研究者的目光纷纷转向新型疫苗和诊断试剂的研制和开发,因此RHDV唯一的结构蛋白-VP60的研究成为热点。国外对RHD基因工程疫苗已进行了广泛的研究,并取得了令人满意的研究成果,在国内只见一篇相关研究的文献[2]报道。
RHDV核衣壳(VP60)由2个同心圆构成,VP60的N端构成核衣壳内层,C端构成核衣壳外层。VP60蛋白的氨基酸由N端到C端又可分A、B、C、D、E、F 6个区域。在T7RNA聚合酶的控制下,西班牙学者E.Viaplana等[3]在大肠杆菌中表达了RHDV衣壳蛋白-VP60的五个重叠部分(A、B、C、D、E)。经过纯化,研究了这些变性蛋白片段的抗原性,用Western blot分析,这些片段能与自然感染和疫苗接种兔的血清起反应。在接种灭活疫苗时,制备的抗血清与片段A和B的反应性约是C、D、E的100倍,在用其它疫苗接种时,片段A、B、C、D的抗原性得到了相似的结果,表明VP60主要抗原表位出现在N末端。VP60氨基末端区域包括连续的抗原决定簇,这些抗原决定簇无论是在病毒感染还是接种灭活疫苗时都能诱导免疫反应。在自然感染动物体内,检测不到抗C片段的抗体,可能是因为在自然感染过程中,该区域在病毒复制过程中被埋在衣壳内部。D段要和杯状病毒VP60向外伸展的P1、P2结构域相结合,这与杯状病毒三维特征相一致。E段与疫苗接种动物的血清和自然感染动物的血清的反应性都较低。研究表明重组RHDV VP60的N末端(A、B段包括最初的175个氨基酸)比VP60的其它部分具有更高的抗原性,这个区域在自然感染和疫苗接种时都具有免疫性并且该部分包括诱导免疫反应的重要抗原决定簇。
本试验用RHDV CD株人工感染家兔,发病死亡后,取肝脏匀浆,用Trizol试剂从匀浆上清中提取病毒RNA。根据Genbank已发表序列保守区设计并合成引物,采用RT-PCR方法扩增了RHDV衣壳蛋白VP60主要抗原表位基因。测定了其核苷酸序列,并与其它13株来自中国(WX-84、NJ、Harbin) 株、法国(99-05、00-13、SD)株、墨西哥Mexico-89株、意大利BS-89 株、英国(Rainham、Wriezen)株、西班牙AST-89株、捷克Crech-V351株、美国Iowa2000株的RHDV VP60同一序列进行了核苷酸及推导氨基酸序列的同源性比较。结果发现,CD株与WX-84株核苷酸序列同源性最高达97.5%,与其它毒株的同源性在92.0%~96.6%之间,其它毒株间的同源性在90.3%~99.4%之间。与WX-84株、墨西哥Mexico-89株氨基酸序列的同源性最高,99.4%,与其它毒株的同源性在96.6%~98.3%之间,其它毒株间的氨基酸同源性在96.0%~100.0%之间。系统发生树中所表明的各毒株间的亲缘关系与动物流行病学资料相符[4]。墨西哥因从中国引入冷冻兔肉而引起RHD的暴发,同年捷克也有该病流行。西班牙与法国株在同一组,可能是由于西班牙和法国在地理位置上相邻所致。有研究报道,RHD在欧洲的暴发是同时从意大利和东欧两个方向席卷欧洲大陆的,因此,意大利与英国株在同一组与动物流行病学资料相吻合。法国株与中国株、美国株同在一个分枝,可能与法国直接从中国输入活兔或兔肉有关。RHDV各分离株VP60主要抗原表位基因核苷酸及推导的氨基酸序列的同源性均在90%以上,显示出高度保守性。RHDV各分离株之间VP60基因的高度保守性是RHDV只有一个血清型的原因所在,也是目前现有RHDV疫苗能够提供有效保护的原因之一[5]。本研究所克隆的VP60主要抗原表位基因,将为RHDV诊断试剂和基因工程疫苗的研制奠定基础。本研究所克隆的片段只是VP60的一部分,该片段是否能代表VP60乃至整个基因组,尚有待进一步研究。
[1]刘怀然,陈洪岩,关云涛,等.兔出血症病毒衣壳蛋白VP60基因克隆与序列分析 [J].中国预防兽医学报,2002,24(5):322-324.
[2]严维巍,崔治中,王永坤.中国株兔出血症病毒衣壳蛋白VP60基因在大肠杆菌中的表达及其免疫原性鉴定[J].中国兽医学报,2003,23(5):447-449.
[3]Viaplana E,Plana J,Villaverde A.Antigenicity of VP60 structural protein of rabbit haemorrhagic disease virus[J].Arch Viol,1997,142(9):1843-1848.
[4]Moss S R,Turner R C,Trout P J,et al.Molecular epidemiology of rabbit haemorrhagic disease virus[J].J Gen Virol,2002,83(10):2461-2467.
[5]严维巍,崔治中,王永坤.兔出血症病毒中国株衣壳蛋白基因的克隆和序列分析[J].中国病毒学,2003,18(2):129-133.
猜你喜欢
解放军医学院学报(2022年1期)2022-04-02
科学大观园(2022年2期)2022-01-23
科学导报(2021年82期)2021-12-17
文萃报·周二版(2021年47期)2021-12-14
生命科学研究(2021年4期)2021-09-08
智慧健康(2021年2期)2021-03-15
温州医科大学学报(2019年4期)2019-04-28
医药前沿(2018年27期)2018-01-16
中国猪业(2017年11期)2017-12-11
中国免疫学杂志(2017年1期)2017-01-17
