
不同年龄杉木林土壤氮矿化及其对施肥增温的响应
2010-03-10 01:47项文化田大伦
湖南农业科学 2010年5期
陈 瑞 ,项文化 ,2,徐 晓 ,田大伦 ,2,刘 佳 ,方 晰 ,2
(1.中南林业科技大学生命科学与技术学院,湖南长沙 410004;2.湖南会同杉木林生态系统国家野外科学观测研究站,湖南长沙 410004;3.湖南省农林工业勘察设计研究总院,湖南 长沙410007)
氮是植物生长中不可缺少的营养元素,但在森林生态系统的土壤中,除氮极端缺乏条件下植物能吸收简单有机氮(DON)外[1],99%以上的氮是不能被植物直接吸收利用,需要经过微生物的矿化作用转化为有效氮才能被植物吸收利用。有效氮一般以铵态氮(NH4-N)、硝态氮(NO3-N)等形态存在[2]。有效氮的供应状况往往影响森林生态系统的生产力,因此森林土壤中氮的矿化过程对生态系统生产力和氮循环起着十分重要的作用[3]。同时,氮的有效性与微生物的活动、植物吸收的过程、氮的固定有着紧密的联系[4]。
国际上普遍采用实验室培养法(Laboratory incubation)来控制土壤温度和水分进而研究土壤氮的矿化过程。温度是影响氮矿化过程中最主要的环境因子之一,施肥也会提高土壤的供氮能力。近年来,国内外学者对温度、水分等环境因子[5-6]、施有机或无机肥对矿化的影响[7]、不同森林类型以及不同土地利用类型土壤氮矿化差异进行了的研究[8-9]。土壤的净矿化速率与温度呈正相关;而净硝化速率在低温段与温度正相关,在高温段则呈负相关,而且整个培养期硝态氮的变化不大[3];落叶林的土壤比针叶林的土壤显示出更高的氮矿化率[10];其中,落叶林土壤的硝化速率很高,而针叶林土壤中却是很低或者几乎没有[11]。
杉木是我国南方主要的用材林树种,不同年龄阶段杉木对土壤养分特别是氮的需求不同[12-13]。在杉木林的生产实践过程中施肥以及未来全球气候变化(如温度升高)等对土壤氮矿化产生怎样的影响,目前还缺乏这些方面的研究。笔者在湖南省会同县选择3个不同年龄的杉木人工林,并用阔叶林为对照,采用实验室培养法控制温度和施肥量,研究其土壤氮矿化速率,为进一步探讨杉木林土壤氮循环和氮供应状况及其对施肥、温度升高的响应提供理论依据。
1 研究地点及样地概况
研究在湖南省会同县进行,属典型的中亚热带湿润性气候,年平均温度为16.8°C,年平均降水量在1 100~1 400 mm之间,年平均相对湿度约为80%。地貌为低山丘陵,地层属于震旦纪板溪系灰绿色变质板页岩,土壤为山地黄壤。
土壤样品来源于3个研究地点,共4块试验地,其中第1、2块分别设立在湖南会同杉木林生态系统国家野外科学观测研究站(26°50′N ,109°45′)的试验基地 II、III集水区内,海拔 270 ~ 400m,II、III集水区内分别为1996年和1988年营造的第2代杉木林。II集水区林分年龄13 a,平均树高9m,胸径8 cm,林分2 490株/hm2。III集水区林分年龄21 a,平均树高13.3 m,胸径16.1 cm,林分密度1 967株/hm2。II、III集水区林下代表性植被有杜茎山、柃木、拔葜、狗脊等。第3块设立在与生态站相距2 km的广坪镇杨家渡村桃子冲(27°50′N,109°45′E),海拔290 ~410 m,坡度15°~30°之间,为 2007 年营造的杉木林,林龄2 a,平均树高为2 m,平均胸径为4 cm。第4块设立在湖南会同鹰嘴界国家自然保护区(26°50′N,109°54′E) 的次生阔叶林内,海拔 330~370m,主要组成树种为拟赤杨、枫香、青冈、南酸枣,林分密度1 596株/hm2,平均胸径10.9 cm,平均树高10.1m,每公顷基径总面积22.18m2。
2 研究方法
2.1 土样采集
在4个不同林地中的山顶、山坡、山脚分别设置3块20 m×30 m小样方,构成3个重复。2009年6月,在4个林地的12个小样方内采集0~10 cm的土样,每3个重复土样充分混合在一个布袋,共有12个,供实验使用。并取一定重量的土样装入铝盒内做含水率测定,同时用环刀法测定土壤容重。采集的土样立即带回实验室冷冻。
2.2 实验处理方案
把12个小样方采集的土样过1 mm的土筛,每个森林类型的一部分混合后分别装入12个250 mL的锥形瓶,每个锥形瓶放入100 g新鲜土样。其中,6 瓶按照 10 g/m2施入硝酸铵(NH4NO3)肥料,另外6瓶不作处理。瓶口用塑料保鲜膜封口,膜上扎若干小眼以通气。然后每一森林类型分别将3瓶对照和3瓶施肥处理放入两个培养箱内培养,其中一个培养箱的温度设定为16.8°C(与试验地年平均气温一致),另一个温度设定为 18.8°C(增加 2°C),培养时间为23 d。剩余部分土样用作理化性质测定。
2.3 样品理化分析
用于测定土壤含水量的土壤样品放入105°C左右烘箱内6 h烘干至恒重,测定土壤含水率。取一部分新鲜土样风干后过筛,用吸管法测定土壤的质地;半微量凯氏法测定全氮含量;重铬酸钾外加热法测定土壤总有机碳(TOC);氢氧化钠熔融法测定土壤全氮和全钾的含量;按土水比为1∶5的比值来测定土壤pH值。培养前后的土壤用酚二磺酸比色法测定NO3-N的含量,氯化钾浸提法测定NH4-N含量。
2.4 数据统计分析
根据培养后与初始值的差值来计算土壤氮净矿化速率,即土壤净矿化速率=(培养后的NH4+-N量-培养前的NH4+-N量)/培养天数;土壤净硝化速率=(培养后的NO3--N量-培养前NO3--N的量)/培养天数。
以土壤pH值、容重、质地、有机碳、NH4+-N、NO3--N、全氮、全钾、C/N为因变量,不同森林类型为自变量,通过方差分析比较不同森林类型之间的差异。以净矿化速率、净硝化速率为因变量,以不同森林类型、不同温度和施用肥料为自变量进行方差分析,比较不同森林类型、增温和施肥之间土壤氮矿化差异。数据统计分析在JMP软件中进行。
3 结果与分析
3.1 4种森林类型的土壤特征比较
阔叶林的pH值、容重、土壤壤粒含量、土壤黏粒含量均要比杉木林的低(表1)。阔叶林的土壤沙粒含量比杉木林的高,差异显著。在不同年龄杉木林中,2 a杉木林土壤的pH值最低。土壤容重、砂粒含量、含水量均随着杉木林年龄的增长呈现递增的趋势,各年龄林分土壤之间差异显著;土壤黏粒却随着杉木年份的增长呈现递减趋势(表1)。
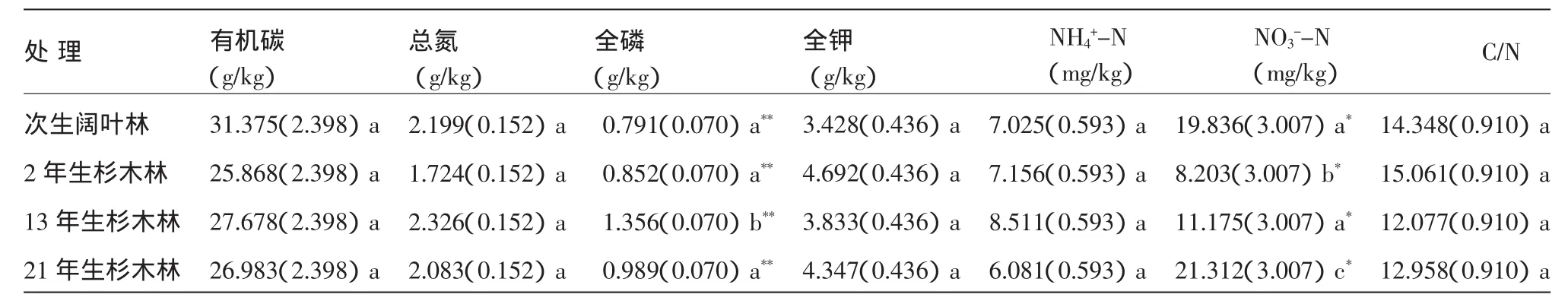
表2 3个不同年龄杉木人工林及阔叶林土壤化学性质比较
阔叶林土壤的有机碳含量比杉木林高(表2),全P含量却比杉木林的低。各林龄的杉木林中13 a杉木林土壤有机碳、全氮、全磷、NH4+-N含量均最高,2 a杉木林土壤全钾和C/N上含量最高(表2);而NO3--N在21 a杉木林土壤中含量最高,不同年龄杉木林NO3--N含量具有显著性(表2)。
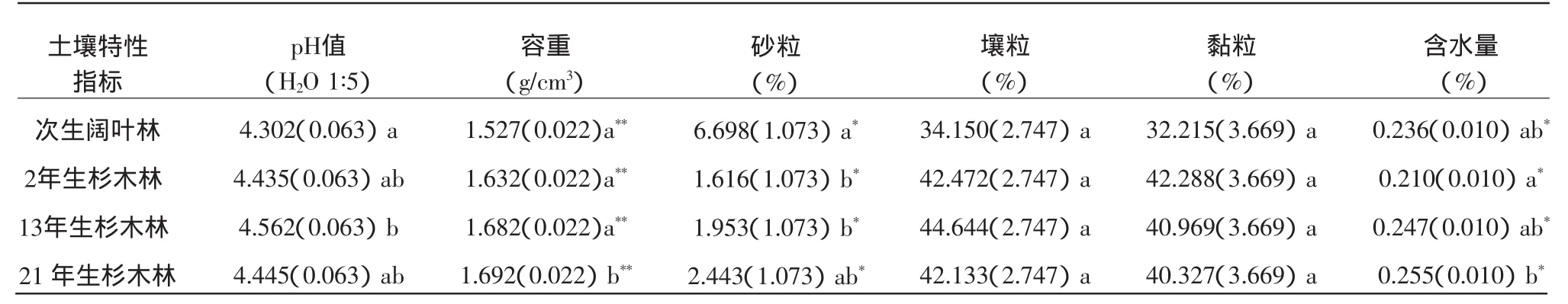
表1 3个不同年龄杉木人工林及阔叶林土壤物理性质比较
3.2 4种不同森林类型土壤氮矿化速率差异

图1 不同林地土壤在不同处理类型下的矿化速率
不同森林类型的矿化速率(p=0.001 7)和硝化速率(p=0.002 3)差异显著。阔叶林土壤净矿化和净硝化速率最低(图1);而在不同年龄杉木林中,21 a杉木林土壤净矿化速率最高,13 a杉木林净矿化速率最低(图1),不同年龄阶段杉木林随着年龄的增长净硝化速率呈递增的趋势,2 a生杉木林净硝化速率最低,21 a杉木林净硝化速率最高(图1)。
3.3 增温和施肥对不同森林类型土壤氮矿化的影响
3.3.1 增温和施肥对不同森林类型土壤净矿化化速率的影响 温度对各森林的土壤净矿化速率影响差异显著(p=0.003 4),在年平均气温16°C基础上增加2°C后,杉木林的净矿化速率下降,表明升温对杉木林土壤净矿化速率有抑制作用,阔叶林的净矿化速率比对照组略有所增加(图1)。施肥对各森林土壤净矿化速率影响没有达到显著水平(p=0.436 2),阔叶林土壤的净矿化速率在施肥后仍然最低,杉木林随着林龄增加,施肥对杉木林土壤净矿化速率的影响增大,21 a的杉木人工林土壤净矿化速率最高(图1)。
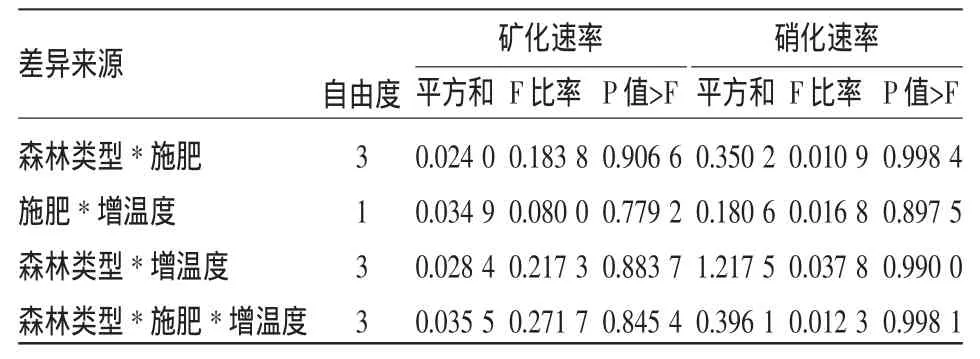
表3 不同因子对不同林地土壤氮矿化、硝化速率影响的方差分析
增温与施肥对各森林土壤净矿化速率的相互作用影响不显著(p=0.779 2),增温和施肥后各森林土壤净矿化速率低于仅施肥后的净矿化速率,但明显高于对照和仅增温的净矿化速率。
3.3.2 增温和施肥对不同森林类型土壤净硝化速率的影响 温度对各森林的土壤净硝化速率影响差异不显著(p=0.201 5),阔叶林土壤净硝化速率最低,随着林龄增长杉木林土壤的净硝化速率呈递增的趋势(图1)。施肥对各森林土壤净矿化速率影响没有达到显著水平(p=0.848 3),但是施肥后阔叶林和杉木土壤净硝化速率明显增加(图1)。
增温与施肥对各森林土壤净硝化速率的相互作用影响不显著(p=0.8975),在13 a杉木林土壤中净硝化速率最高,增加也最快;21 a杉木林施肥同时增温条件下净硝化速率与仅施肥条件下的没有差异(图1)。增温与施肥后,各森林的土壤净硝化速率总体上比施肥组的高,说明温度和施肥对硝化反应起着促进作用(图1)。
4 结论与讨论
温度对土壤氮矿化速率的影响,主要是通过改变微生物的生化过程和耗氧量[14],氮矿化一般随温度而升高[3]。由于次生阔叶林土壤的含水率本身就比杉木林土壤低,因此即便温度升高,微生物在升温但是水分不够的情况下,活跃度变化不大。而不同年龄杉木林土壤中的微生物在升温后,开始活跃,显示出温度对不同森林土壤净矿化速率影响差异显著(p=0.003 4)。有研究表明温度从17°C上升到30°C,氨态氮将逐渐提高,本试验温度还没有增加到达到影响的范围是导致净矿化速率降低的原因之一。硝化细菌在酸性土壤上,硝化反应受到抑制,升温也不能对其起到促进作用,因此温度对各森林的土壤净硝化速率影响差异不显著(p=0.201 5)。
施肥后,土壤中有机碳含量相对下降,由于土壤微生物矿化作用大于同化作用,容易造成氮素损失,降低了土壤中C/N比,使得土壤净矿化速率加快。而在有充足氧气的供应下,土壤微生物氮的矿化作用大于自身吸收。土壤氮矿化受到pH值以及砂砾比例影响大于受到微生物的作用,从而使得施肥对不同森林土壤净矿化速率和净硝化速率影响没有达到显著水平。
土壤无机氮的动态变化是土壤有机氮的矿化作用、土壤淋溶作用、植物根系的吸收相互作用的结果[9],同时受环境条件和土壤特征的影响。研究通过试验控制将土壤单独放在锥形瓶中培养,温度和土壤水分等试验条件相同,还阻隔了雨水淋溶以及植物根系的吸收,排除了环境因子对氮矿化的影响。因此,各森林土壤特征如土壤的pH、含水率、C/N和土壤微生物的不同是影响森林土壤氮矿化的主要原因。
[1] Schimel J P,Bennett.Nitrogen mineralization:challenges of a changing paradigm[J].Ecology,2004,85(3):591-602.
[2] Chapin III F.S.Principles of terrestrial ecosystem ecology[J].New York:Springer-Verlag,2002.202-210.
[3] 周才平,欧阳华.长白山两种主要林型下土壤氮矿化速率与温度的关系[J].生态学报,2001,21(9):1469-1473.
[4] Groffman PM ,Zak D R,Christensen S.et al.Early spring nitrogen dynamics in a temperate forest landscape[J].Ecology,1993,(74):1579-1585.
[5] 陈伏生,余甘露,刘 义,等.温度、水分和森林演替对中亚热带丘陵红壤氮素矿化影响的模拟实验 [J].应用生态学报,2009,20(7):1529-1535.
[6] 李贵才,韩兴国,黄建辉,等.森林生态系统土壤氮矿化影响因素研究进展[J].生态学报,2001,21(7):1187-1195.
[7] 鲁彩艳,牛明芬,陈 欣,等.不同施肥制度培育土壤氮矿化势与供氮潜力[J].辽宁工程技术大学学报,2007,26(5):773-775.
[8] 陈祥伟,陈立新,刘伟琦.不同森林类型土壤氮矿化的研究[J].东北林业大学学报,1999,27(1):5-9.
[9] 严德翼,周建斌,邱桃玉,等.黄土区不同土壤类型及土地利用方式对土壤氮素矿化作用的影响[J].西北农林科技大学学报,2007,(10):103-109.
[10]Cote L,Brown S,Pare D,et al.Dynamics of carbon acid nitrogen mineralization in relation to stand type,stand age and soil texture in the boreal mixedwood[J].Soil Biology and Biochemistry,2002,(32):1079-1090.
[11]Ste-Marie C and Pare D.Soil,PH and N availability effects on net nitrification in the forest floors of a range of boreal forest stands[J].Soil Biol Bilchem,1999,(31):1579-1589.
[12]向华升,高 人,马红亮.不同年龄杉木人工林土壤无机氮比较研究[J].亚热带资源与环境学报,2008,3(1):24-30.
[13]Wang L,Wang J,Huang J.Net Nitrogen Mineralization and Nitrification in Three Subtropical Forests of Southwestern China[J].Dynamic Soil,Dynamic Plant,2008,2(1):33-40.
[14]Sierra J.Temperature and soilmoisture dependence of N mineralization in intact soil cores[J].Soil Biology and Biochemistry,1997,(29):1557-1563.
猜你喜欢
北京大学学报(自然科学版)(2022年4期)2022-08-18
草业学报(2021年9期)2021-09-22
农民致富之友(2020年8期)2020-05-11
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
农业环境科学学报(2017年2期)2017-03-20
广西林业科学(2016年2期)2016-03-20
亚热带资源与环境学报(2015年4期)2015-01-22
东北师大学报(自然科学版)(2014年1期)2014-02-27
亚热带资源与环境学报(2014年3期)2014-01-23
