
植物间水平基因转移
——基因交流新途径及其农业利用潜力
2024-03-23 09:17王挺进张爱珺李旭青道丽筠陈利萍
浙江农业学报 2024年2期
杨 洋, 袁 璐, 刘 彬, 王挺进, 张爱珺, 刘 柯, 李旭青, 道丽筠, 袁 鑫,陈利萍,,*
(1.浙江大学 农业与生物技术学院 园艺系,浙江 杭州 310058; 2.青海省乌兰县农牧和乡村振兴局,青海 乌兰 817199; 3.浙江大学山东(临沂)现代农业研究院,山东 临沂 276000)
水平基因转移(horizontal gene transfer, HGT),又称横向基因转移(lateral gene transfer, LGT),是指通过受精以外的方式进行的跨物种遗传物质传递,对生物进化有着十分重要的意义。水平基因转移现象首先于1928年在小鼠的肺炎球菌中被观察到,随后在大肠埃希菌中得到证实[1-3],并在此后成为研究原核生物进化的重要方向。随着二代测序技术的发展和研究的不断深入,研究者们逐渐在真核生物,包括真菌[4-5]、动物[6-7]、植物[8-11]中都发现了水平基因转移现象。目前,尽管人们对生物中水平基因转移的研究方法和发生规模仍有争议,但它在生物进化过程中发挥的作用仍受到了广泛的关注和越来越多的认可。
植物是地球生命至关重要的组成部分,不仅为我们提供了生命活动所必需的氧气、食物等资源,也具有着重要的观赏和研究价值,更是我们了解物种起源和物种进化的重要材料。研究表明,水平基因转移现象在植物的进化过程中持续发生,并帮助植物向环境适应方向不断地进化。早在1970年,研究者就利用抗性培养基筛选的方法证明了蓝藻中基因交换现象的存在[12]。但直到进入21世纪后,随着测序技术的发展和系统发育分析法的应用,大量植物间水平基因转移的现象才得以发现。2003年,相继报道了两例对高等植物间水平基因转移的研究[13-14]。随后的研究进一步发现,植物的细胞核和细胞器基因组均可以进行转移。水平基因转移现象不仅能够在植物进化过程中自然发生,也可以通过嫁接等方法人工操作诱导产生,从而赋予植物新的性状[15]。因此,对植物水平基因转移现象的探究,不仅有利于了解植物的进化过程,也可以为植物新性状的起源与形成提供确凿证据。本文综述了一些典型的植物间水平基因转移事件,以及近年来国内外最新研究进展,以期为植物水平基因转移在农业生产上的利用提供理论依据与新途径。
1 植物间水平基因转移的途径
一般而言,水平基因转移需要供体与受体间的直接接触,这种情况往往发生在寄生植物与其寄主之间或嫁接植物的接穗和砧木间。除此之外,植物还可以通过媒介进行基因的水平转移。因此,对植物而言,水平基因转移可以通过直接和间接两种途径来进行。其中,直接转移又包括了寄生和嫁接两种方式,即寄生介导的水平基因转移与嫁接介导的水平基因转移。
1.1 寄生介导的水平基因转移
在目前的报道中,大部分水平基因转移现象都是在寄生植物和寄主之间产生的。许多寄生植物能够通过吸器与其寄主产生细胞间的紧密连接,这可能是两者间频繁基因交流的基础。有研究称,随着异养程度的增强,植物通过水平转移获得的基因数量也会上升[16]。因此,用寄生植物来探究水平基因转移现象及其发生规律具有重要意义。
寄生性杂草往往会为农业生产带来巨大的危害。多项研究表明,寄生植物通过水平转移获得的基因能够在其寄生性中起作用。列当科为唇形目的重要类群,其下全部为草本寄生植物,包括15个属的150多种植物,我国拥有其中的9属[17]。在列当科的这些寄生植物中,存在着一些农业杂草,如寄生在禾本科农作物的独脚金(Strigaasiatica)以及寄生在豆科、菊科、茄科以及葫芦科作物上的列当(Orobanchecoerulescens)。研究发现,列当科植物与其寄主间存在着大量的水平基因转移现象。Yang等[16]对该科的3种寄生植物变色直果草(Triphysariaversicolor)、黄独脚金(Strigahermonthica)以及分枝列当(Phelipancheaegyptiaca)进行系统发育分析后,发现了52个高置信度的水平基因转移事件,其中多数基因优先在吸器中表达,并有可能在这些植物与寄主的相互作用中发挥功能。随后,Kado等[18]对列当科的另外5种寄生植物进行基因组测序,并对其豆科、禾本科寄主的参考基因组和转录组进行联合分析后,在两种专性寄生植物小列当(Orobancheminor)和野菰(Aeginetiaindica)中共发现了106个水平转移的基因,它们主要富集在非编码RNA(ncRNA)代谢和维生素代谢中。此外,为进一步加强对独脚金寄生性的理解,Yoshida等[19]对其进行了基因组测序,也发现了34个水平转移的候选基因。
菟丝子(Cuscuta)同样是一种对农业有严重危害的恶性寄生杂草,能够寄生在豆科、葫芦科、茄科、十字花科的多种作物上,导致作物减产甚至死亡。研究发现,菟丝子与其寄主间也存在着广泛的水平基因转移,包括由菟丝子到寄主[20-21],以及由寄主到菟丝子的转移[22-24]。为了探究菟丝子中水平基因转移的规模,Yang等[25]对原野菟丝子(Cuscutacampestris)进行了从头转录组装和系统发育分析,鉴定到了108个能够转录并且可能具有功能的水平基因转移事件,它们富集在吸器表达,在防御反应和氨基酸代谢途径中发挥作用,可能有助于菟丝子与寄主的相互作用。随后,对原野菟丝子的核基因组测序也发现了来自至少32种不同供体的64个水平转移的候选基因,其中大部分来自其豆目或石竹目寄主。它们中的部分在吸器中的表达明显上调。该研究再次证明,菟丝子通过水平转移获得的基因可能在寄生过程中发挥作用[23]。吸器是寄生植物入侵寄主以及从寄主中获取营养物质的器官,对寄生植物的生长发育具有重要意义。因此,对寄生植物与寄主间水平基因转移现象的探究,可能有助于对寄生植物的了解以及农业生产中对寄生性杂草的防治。
除了这两类农业杂草外,还有一些寄生植物及其寄主中也存在着水平基因转移现象,如大花草科植物及其葡萄科寄主[26-28],列当科寄生植物肉苁蓉(Cistanchedeserticola)及其寄主梭梭(Haloxylonammodendron)[29],以及锁阳(Cynomorium)及其无患子目、石竹目寄主[30-31]等。总之,这些水平转移基因的发现,对于探究寄生过程的进行以及潜在的农业杂草防治手段都具有十分重要的意义。
1.2 嫁接介导的水平基因转移
由于嫁接能够导致接穗与砧木间细胞的连接,所以嫁接介导基因水平转移是受人工操作控制的方法。从报道的资料来看,对嫁接介导水平基因转移的研究大多在茄科烟草属或种间进行,一般思路为,将嫁接部位放置在抗性培养基上,并进行愈伤组织的诱导以及成功转移材料的筛选(图1)。嫁接之后的水平基因转移现象,最早便是通过这种方法在烟草中观察到的[15]。随后再次通过烟草的嫁接证明了叶绿体基因组在接穗和砧木间的转移,为细胞器在细胞间的运动提供了证据[32-33]。当然,线粒体和核基因同样也可以通过嫁接发生水平转移,且不同基因组的转移可以同时存在[34-35]。
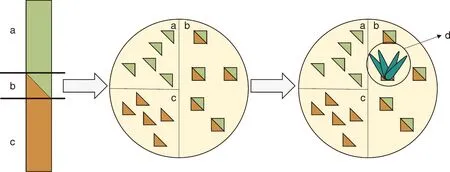
图片参考自Stegemann等[15]的文章。a,接穗;b,嫁接部位;c,砧木;d,再生芽。The picture is taken from the article of Stegemann et al.[15]. a, Scion; b, Graft region; c, Rootstock; d, Regenerating shoots.
上述实验的成功使得嫁接成为研究水平基因转移机制的重要手段。通过对烟草的嫁接和嫁接部位的实时成像观察发现,在伤口的愈伤组织增殖过程中,质体会脱分化成为小的、高度运动的变形细胞器。同时,细胞间形成新的连通孔隙,脱分化的质体通过这些孔隙进入相邻的细胞。这是首次利用嫁接技术获得质体在细胞间水平转移的可视化证据[36]。
1.3 媒介介导的水平基因转移
一般认为,水平基因转移的实现需要细胞间的直接接触,但仍有部分研究发现,在远缘且无直接接触关系的物种间也存在水平基因转移现象。因此,研究者们提出了载体介导假说,即遗传物质能够以细菌、病毒等载体为媒介,在不同植物间转移。
病毒是水平基因转移的理想载体,它具有强大的捕获和整合外源基因的能力,通过对其他生物体的感染,可以将这些捕获到的基因再次转移[37]。在病毒介导下,水平转移的基因可能来自线虫、昆虫等多种生物,但目前对病毒介导的植物间基因转移的报道很少。首次通过实验验证的,是伊朗甜菜曲顶病毒(BeetcurlytopIranvirus, BCTIV)在感染甜菜(Betavulgaris)后能够产生病毒/寄主杂交DNA分子,并与病毒同时在其他受感染的植物中传播[38]。
错位授粉假说同样是水平基因转移的假设机制之一。错位授粉假说是指供体的花粉粒在与其有生殖系统隔离的受体物种的柱头上萌发,随后一些外源基因与受体的基因组进行重组和整合,并在选择中遗传给后代的水平基因转移机制,一般在亲缘关系较近的物种之间发生。目前这种现象在禾本科的毛颖草属(Alloteropsis)与C4植物、狗尾草属(Setaria)与水稻(Oryzasativa)以及早熟禾属(Poa)与羊茅(Festucaovina)间均有推测报道,但尚未通过实验证实[39-41]。
此外,水平基因转移的可能载体还包括细菌、真菌、昆虫、线虫、转座子等,但目前对媒介介导的水平基因转移现象的研究,还并不充分。
2 植物中水平转移基因的分类
植物细胞中含有3个相对独立的基因组,分别是核基因组、叶绿体(质体)基因组以及线粒体基因组,每种基因组都可能成为基因水平转移的供体或受体。当然,不同基因组发生交换的频率不同,也会有基因二次转移现象的发生,如基因可能会先从寄主的叶绿体转移到细胞核,然后再转移到寄生植物中[42]。此外,虽然大多数转移的都是DNA片段,但RNA片段的转移也同样存在[43]。
2.1 线粒体基因转移
线粒体是细胞代谢的中枢,在细胞的生长分化、能量供应与凋亡等生理活动中发挥着重要功能。它被认为是由水平基因转移形成的,来源于一种内共生的α-蛋白杆菌[44]。对24种植物进行研究后发现,线粒体在细胞内不同基因组间的转移非常普遍,它既可以作为转移序列的受体,也可以作为供体[45]。与叶绿体和核基因组相比,线粒体更加活跃,更容易发生水平基因转移。植物的线粒体能够输入DNA或RNA,并容易发生融合;线粒体也有一套活跃的同源重组系统,这些特性都可能帮助线粒体参与到基因的水平转移中[46-47]。在某些情况下,植物通过水平转移获得的线粒体基因甚至能够占据其蛋白质编码基因含量的80%[48-51]。此外,线粒体也会接受来自细胞核和叶绿体的基因[52-54]。因此,线粒体基因组可以被看作是通过基因转移整合而成的基因组。
线粒体是为生命活动提供能量的重要场所,其基因组中包含一些与能量代谢相关的基因。其中,cox1编码细胞色素氧化酶亚基1(cytochrome oxidase subunit 1),在氧化磷酸化中发挥作用[55]。大量研究表明,cox1基因中的I组内含子在不同植物中经历了多次水平转移,其发生规模和频率甚至可以用猖獗来形容。它是第一例在植物中得到证实的线粒体水平转移基因,最初由被子植物通过真菌的转移获得,随后在不同被子植物间又进行了多达约70次独立的转移[56-58]。内含子介导的调节是真核生物基因表达调控的重要机制。因此,不同植物cox1基因中I组内含子在不同位置的插入,可能会对该基因的表达起到意想不到的作用。此外,cox1基因本身也可以在不同物种间进行转移。藤本寄生植物无根藤(Cassythafiliformis)的线粒体基因cox1 具有两种不同起源的拷贝,其中一种可能来自木兰类植物的垂直遗传,并且具有完整的开放阅读框(open reading frames, ORFs),而另一种则为假基因拷贝,它可能来自旋花科植物的水平转移[59]。
除cox1外,线粒体水平基因转移中报道较多的还包括NADH脱氢酶基因nad1和nad5[14,60],核糖体蛋白基因rps2和rps11[13,54,61],以及ATP合成酶基因atp1和atp8[13,20,31,62]等(表1),它们均是调控线粒体生物氧化作用的重要基因,其转移在低等植物和高等植物中普遍存在。研究发现,硅藻中的cox1_g2_B内含子可能通过水平转移进入了其他藻类[63];蕨类植物蕨萁(Botrychiumvirginianum)的线粒体中存在着来自被子植物的nad1B-C和matR片段[64];裸子植物北非雪松(Cedrusatlantica)的线粒体中也含有可能来自被子植物的rps3和rpl16序列[65]。线粒体基因水平转移规模之大的另一个表现是,这种转移甚至可以双向发生。早前的研究证明,菟丝子通过水平转移将atp1转移到了其寄主车前属(Plantago)中,并在寄主中变为了假基因[20];近期对菟丝子线粒体基因组进行分析后发现,其Pachystigma亚属的atp1基因同样是通过水平转移从唇形目寄主中获得的,并可能已经对其原始基因进行了功能性替代[66]。最具代表性的是裸花蛇菰(Lophophytummirabile),它从豆科寄主中获得了大量的线粒体基因序列,且几乎所有的这些外来基因都与其原始基因构成了嵌合基因,或取代了原始基因。其中,有23个外源基因可以转录为mRNA,说明它们极有可能被翻译为蛋白质,并在呼吸作用中发挥功能[48-49,67]。值得一提的是,裸花蛇菰核基因组中同样存在来自豆科寄主的水平转移基因sdh3,它能够与线粒体蛋白相互作用[68]。这项研究说明了一个有趣的现象,即外源基因在新的物种中也能够继续合作。
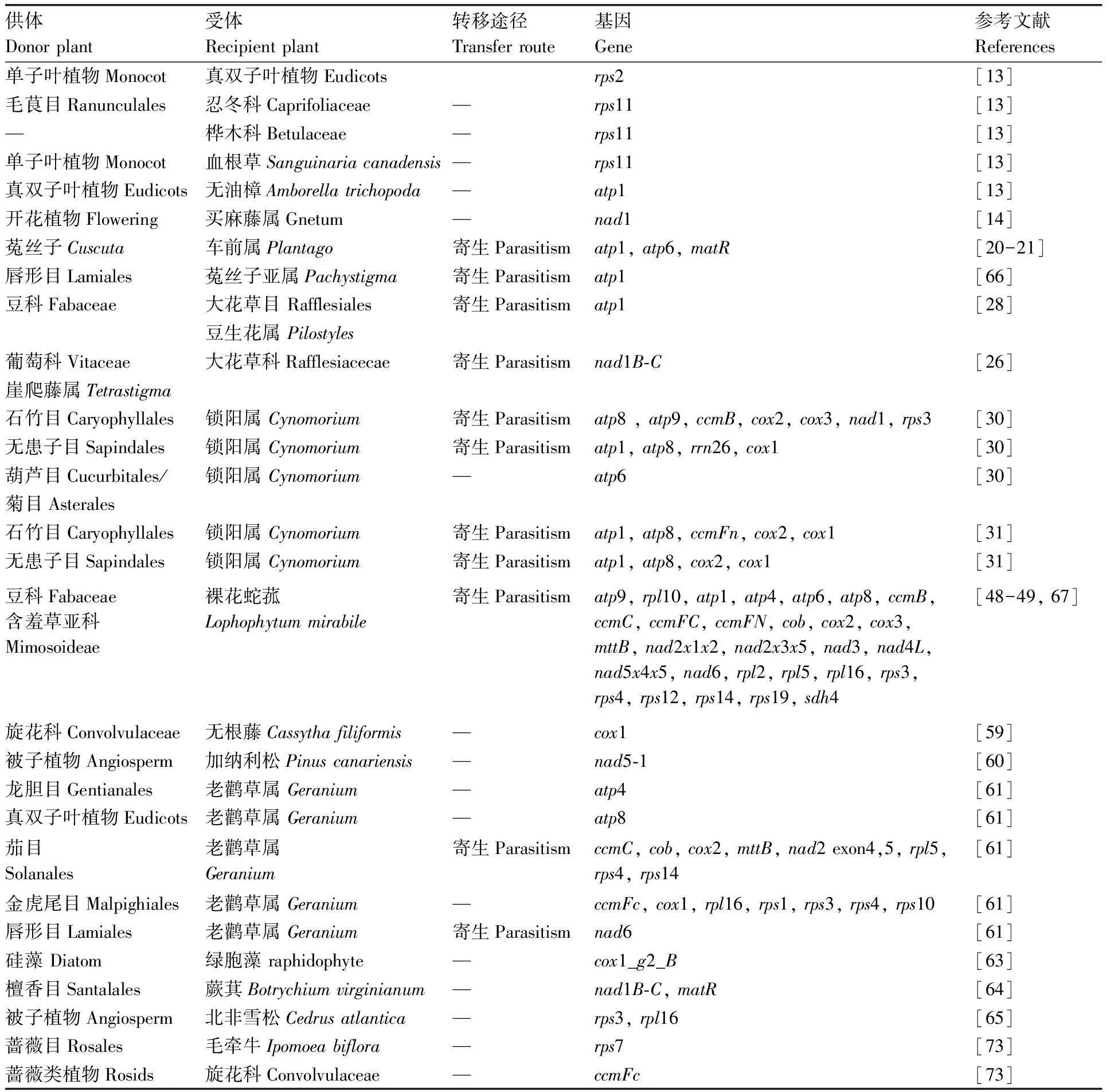
表1 植物间线粒体基因的水平转移
线粒体对植物育性的控制也是一个备受关注的热点。细胞质雄性不育(cytoplasmic male sterility, CMS)是指细胞核不育基因和细胞质不育基因共同控制的雄性不育,其表现主要为无法产生功能性花粉[69]。Woloszynska等[70]发现,一些菜豆属(Phaseolus)物种的线粒体基因组中,控制雄性不育的片段pvs(Phaseolusvulgarissterility sequence)中包含了叶绿体trnA内含子序列,称为pvs-trnA,它们可能来源于非双子叶植物的水平转移。随后,Gurdon等[34]将细胞质雄性不育的烟草与育性正常的烟草进行嫁接后,发现线粒体DNA能够在接穗和砧木间移动,并恢复烟草的育性。同时,他们还挖掘了与胞质雄性不育相关的基因orf293,并确认了它在可育系中的存在和不育系中的缺失。由此可见,水平基因转移不仅有助于细胞质不育系与可育系的构建,并且可用于CMS相关基因的鉴定,对于育种工作有重要的意义。
但并非所有物种的线粒体中都存在大量的水平基因转移现象。研究发现,槲寄生(Viscumscurruloideum)的线粒体中几乎不存在水平转移的基因[71]。同样,菟丝子线粒体基因组中水平转移基因的比例也远低于此前的预测[72-73]。这种情况出现的原因,可能是这两个物种有丝分裂的基因组较小,限制了对外来序列的整合能力。与之相反的例子是无油樟(Amborellatrichopoda),它从其他陆地植物中获得了20个线粒体蛋白基因的拷贝,且该植物的线粒体基因组包含了6个基因组大小的外源线粒体DNA,它们来自绿藻、苔藓以及其他的被子植物,其中甚至包括3种绿藻和1种苔藓的完整基因组[74-76]。
2.2 叶绿体基因转移
叶绿体是植物光合作用的场所,它同样来源于一种典型的水平基因转移现象,最初由一种内共生的蓝细菌形成[44]。研究发现,质体通常不会发生融合和重组,因此叶绿体大多是以整个细胞器为单位进行转移的[32]。细胞器在不同物种间的转移可能会导致核质不亲和(nucleocytoplasmic incompatibility)现象的发生,因此这可能也是阻碍物种之间,尤其是远缘物种间质体水平基因转移的因素之一[32]。
叶绿体及其片段的转移多发生在叶绿体与细菌、真菌间或植物细胞的内部,且出现在植物进化的早期[77-78],而涉及植物间叶绿体水平基因转移的报道并不多,其中最早的研究来自一组寄生性的非光合开花植物。对小苞列当属(Phelipanche)中几种植物的叶绿体进行系统发育分析后,发现了一个包含rps2、trnL-F和rbcL的水平基因转移区域,且这段转移片段的供体也是寄生植物[79]。通过对寄生植物肉苁蓉质体基因组的分析,发现它从寄主梭梭中获得了rpoC2基因的1个拷贝[29]。对野菰的叶绿体基因组进行组装后发现,其质体基因组中的atpH基因是从另一个被子植物的质体中转移过来的[80]。此外,葡萄(Vitisvinifera)的线粒体基因组中存在大量与野胡萝卜(Daucuscarota)叶绿体基因组高度相似的序列,且这些序列与其他已发表的叶绿体基因组序列没有同源性,表明这可能是第一例发现的由线粒体向叶绿体进行水平基因转移的事件[81]。但总体来说,叶绿体中外来序列的天然插入现象是相对少见的(表2)。

表2 植物间叶绿体基因的水平转移
然而,与进化过程中自然发生的频率相反,在目前报道的嫁接介导的水平基因转移中,对叶绿体转移的研究是最多的[15,32-33,82]。这一现象产生的原因,除了叶绿体在水平基因转移的研究上存在的天然优势(如通常不会融合和重组,报告基因能够稳定存在并表达)外,还有可能是嫁接时多采用亲缘关系较近的物种,从而在一定程度上避免了核质不亲和现象的发生[32,36]。
此外,对叶绿体水平基因转移的研究中也存在一些有趣的报道。Hedenäs等[83]分别在受石灰干扰的沼泽(代表初生演替栖息地)和天然沼泽(代表演替后期)中采集了大叶镰刀藓(Scorpidiumcossonii),并在前者的质体中发现了来自拟三列真藓(Bryumpseudotriquetrum)的rpl16片段,而后者质体中不存在该片段,说明该转移事件可能发生在近几十年,即其自然栖息地受到严重干扰后的这段时期。这项研究为水平基因转移的发生原因提供了新的思路,即水平基因转移可能会在植物的生存环境受到剧烈改变时发生。
2.3 核基因转移
细胞核是细胞遗传以及各种生理活动的控制中心,它可以作为基因转移的受体,在进化过程中整合许多其他物种以及细胞器的基因。比如,叶绿体和线粒体在进化中丢弃的一部分基因便通过内共生转移到了核基因组中[84]。鉴于细胞核对生命活动的主导作用,其基因组的变化必然会对植物产生更加深远的影响。对植物间核基因的转移多有研究[22,27,43,85],尽管大部分转移的片段已失去作用或功能未知,但仍有一部分水平转移的基因已被证明在受体植物中展示出了作用(表3)。
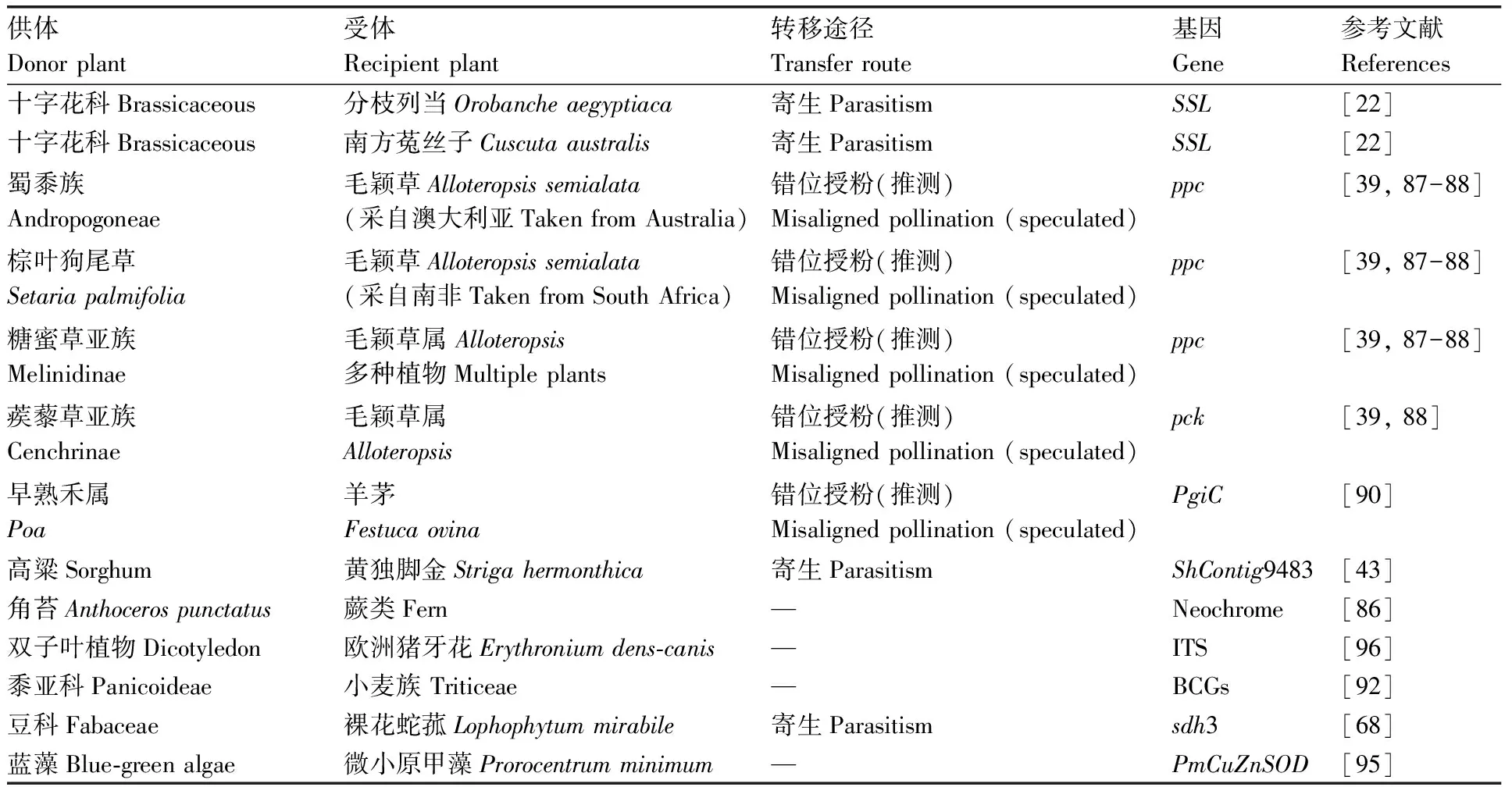
表3 植物间细胞核基因的水平转移
水平基因转移对植物光合作用机制的演变有着十分重要的作用。蕨类植物和C4植物都具有独特的光合能力,它们分别能够适应弱光和提高CO2利用效率。Li等[86]发现,蕨类通过水平转移从角苔(Anthocerospunctatus)中获得了红/蓝光嵌合光受体(neochrome)编码基因,并因此获得了在弱光下茁壮成长的能力,对其进化起到了非常重要的作用。同样,Christin等[39]通过对禾本科毛颖草属C4植物中编码C4循环关键酶的基因ppc和pck进行分析后发现,它们可能是通过多次水平转移得到的。随后的基因组测序证明了这一观点,即毛颖草属中不同植物的ppc是通过3次水平转移分别获得的,而pck则由它们的共同祖先获得[87-88]。对这些基因的酶促反应动力学实验证明,与毛颖草属中自然进化的ppc比,3个水平转移的ppc基因催化效率都显著提高,说明它们可能在C4光合途径中发挥重要作用[89]。
除毛颖草属外,有更多发生在禾本科的水平基因转移现象已被报道。禾本科植物中包含大量草本植物,它们多样的生长方式,广泛的地理分布以及丰富的基因组资源为水平基因转移提供了良好的研究条件。研究发现,羊茅从早熟禾属中获得了编码胞质磷酸葡萄糖异构酶(cytosolic phosphoglucose isomerase, PgiC)基因的一个额外拷贝,并与自身的PgiC基因一起,对其环境适应性起到了作用[90]。常见作物中也存在着大量水平基因转移。比如,Hibdige等[91]分别在玉米和小麦中检索到了11和10个HGT事件,它们可能与作物的抗性有关。更加明确的例子是,小麦族的祖先通过水平转移从黍亚科的祖先获得古老的Bx生物合成基因簇(BGCs),并在不同染色体上分离成多个片段,通过复杂的进化过程得以保留,产生苯并噁唑嗪酮类化合物,在植物的抗性中起到了重要作用[92]。此外,种子植物间转座子的水平基因转移现象也是在禾本科中最先发现的,第一例报道是一个MULE转座子在狗尾草属和水稻间的序列相似性[40]。随后对几个谷类物种全基因组测序研究,发现了LTR反转录转座子Route66可以在黍亚科的高粱属(Sorghum)和稻属(Oryza)的几个物种之间转移[93]。但是,不同家族转座子的水平转移能力也不完全相同。导致这种现象的原因,可能与部分转座子进化出了减少依赖特定寄主的能力,或转座子中间体的性质有关[94]。禾本科中存在着许多重要的粮食作物,探究它们水平基因转移发生的规模、频率以及机制,可能会对这些作物的改良起到额外的帮助。
此外,微小原甲藻(Prorocentrumminimum)可能通过水平基因转移或内共生从蓝藻中获得了超氧化物歧化酶基因PmCuZnSOD,并在氧化应激中发挥作用[95]。单子叶植物欧洲猪牙花(Erythroniumdenscanis)中含有来自双子叶植物的核糖体DNA内部间隔转录区(internal transcribed spacer, ITS)序列,证明双子叶植物和单子叶植物之间也可以发生核基因的水平转移[96]。而前文所述的寄生植物从寄主中获得的功能性基因,也有许多是定位于细胞核中的[16,22-25]。这些研究说明,植物通过水平转移获得的核基因,能够在各种生命活动中发挥重要的作用。
3 水平基因转移在农业中的应用潜力
植物间的水平基因转移现象涉及细胞核、叶绿体和线粒体DNA。这些转移往往是通过寄生、嫁接以及各种媒介来实现的,并且可能赋予植物体新的性状(图2)。大量研究表明,水平基因转移不仅对生物进化有重要意义,而且有望应用在农业中,为农业生产提供帮助。比如,农杆菌(Agrobacterium)无疑是植物间水平基因转移中最常用的载体,它拥有可以转化并整合到植物基因组上的Ti或Ri质粒,可以在自然状态下感染大多数双子叶植物。利用它的这个特点,人们创造性地开发了植物转基因技术,并广泛用于科学研究。目前农杆菌介导的水平基因转移在作物改良中多有应用,相关综述也层出不穷[97-99],因此本文不做重点阐述。由于植物间的水平基因转移提供了物种之间基因交流的机会,这为农作物性状改良提供了一条新途径,具有重要的农业生产应用潜力。
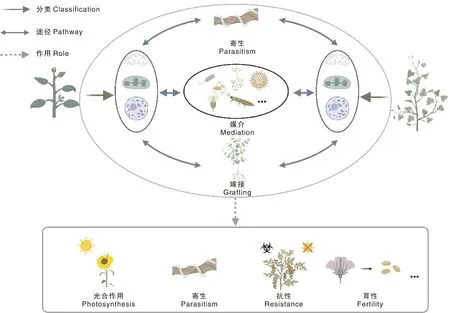
植物能够通过寄生、嫁接以及媒介介导的方式,将线粒体、叶绿体和核基因组的DNA水平转移到其他植物中。这些水平转移的基因可能对受体的光合作用、寄生、抗性及育性等性状产生影响。图片创作自BioRender。Plants can transfer DNA of mitochondria, chloroplast and nuclear genomes to other plants through parasitism, grafting and vector mediation. These horizontally transferred genes may affect the photosynthesis, parasitism, resistance and fertility of receptor. The image was created with BioRender.com.
3.1 水平基因转移与农业杂草的防治
许多寄生植物都从寄主中获得了大量水平转移的基因,它们可能在其寄生性中起到重要的作用[16,18-19,23,25]。虽然目前对于这些转移的基因并没有进行直接的功能研究,但对于它们在寄生过程中的调控作用已有研究报道。作为一种农业恶性杂草,菟丝子的寄生性及其防治受到了许多研究者的关注。Shahid等[100]的研究表明,原野菟丝子在寄生拟南芥和烟草的过程中,在吸器中积累了大量的miRNA,它们能够靶向寄主编码防御蛋白的mRNA并减少其积累,从而对寄生过程起到积极作用。鉴于菟丝子与其寄主间频繁的基因交流,有一种可能是,这些跨物种作用的miRNA起源于通过水平转移获得的基因[101]。随后的研究证明了这一点,即菟丝子中的部分miRNA序列与水平转移的序列高度相似,说明这些序列可能在转移之后进行了几次突变和插入,最后产生了这些miRNA[25,102]。此外,Yang等[25]在研究中发现,有18个菟丝子和列当科的水平转移基因事件是从相同的基因家族获得的,说明植物中水平转移基因的保留可能具有非随机性和趋同性。因此,探究寄生植物与寄主间的水平基因转移现象,可能对寄生性杂草的防治具有普适意义。
3.2 嫁接介导的水平基因转移以及新种质的创制
嫁接是一项传统的农业技术,最初用于多年生木本植物,后来被广泛应用于各种作物中,目前已经拥有超过2 000年的历史。与传统杂交育种相比,嫁接能够快速改良植株性状,因此受到了生产者们的青睐。作为一种无性繁殖的方式,嫁接是一种栽培技术。然而,嫁接诱导的植物间的水平基因转移,可以为农作物新种质的创制提供一种育种新方法。Lu等[82]通过嫁接,将含有虾青素合成操纵子的叶绿体基因组从可以高效生产虾青素的转基因烟草(Nicotianatabacum)转移到了难以进行质体遗传转化的光烟草(Nicotianaglauca)中,并成功在光烟草中诱导了与转基因烟草相似的类胡萝卜素代谢变化。然而,这种方法仍需要传统的嫁接手段,具有一定的物种局限性。因此,Sidorov等[103]创造性地开发出了一种“细胞嫁接”技术,即将两种不同物种的愈伤组织分别损伤后,再混合到一起进行培养。目前,已经成功用这种方法实现了不同转基因烟草间细胞质和细胞核基因组的交流,为作物改良提供了有力的工具。此外,Notaguchi等[104]发现,在烟草中发现的β-1,4-葡聚糖酶能够促进植物嫁接中细胞间的黏附,有利于远缘物种间的嫁接。这些研究拓展了嫁接技术的利用范围,为植物远缘物种间的水平转移奠定了基础。
与转基因育种相比,嫁接介导的水平基因转移同样可能在新种质创制中有独特的优势。比如,许多科学家希望能够在C3植物中完成C4途径的改造,从而培养高产高效的作物。2017年,C4水稻项目(https://c4rice.com)的研究者们通过转基因技术,初步实现了水稻叶片解剖学结构向C4植物的改变[105]。随后,他们将C4植物玉米光合途径中5种酶的编码基因组合到一起,并转化至水稻中,实现了C4途径的部分运行[106]。然而,总体来说,人工创制C4植物的探索进程是非常缓慢的。同时,鉴于目前对转基因作物的应用限制,这些改造作物能否被顺利投放至市场,也是一个未知的问题。研究表明,毛颖草属C4植物中C4途径的两个关键酶基因ppc和pck是通过水平基因转移从其他C4植物中得到的[39]。与其自身进化出的相关功能性基因相比,这些酶的活性更高,说明水平基因转移可能是毛颖草属植物获得C4光合能力的“捷径”[89]。此外,水平基因转移是不同物种间遗传物质的自然传递,与转基因技术相比,其安全性更受研究者与大众的认可。因此,如果选用合适的C3和C4植物,并利用嫁接技术进行水平基因转移的诱导,可能对于C4植物的人工创制有重要的推进作用。
但是,目前对于通过嫁接转移的基因能否在受体植物及其后代中稳定存在的问题,仍需进一步探究。有研究者发现,通过嫁接转移获得的叶绿体和细胞核,能够传递至有性繁殖产生的后代中[32,35],并有望通过基因组适应和选择后形成新种质。但也有研究发现,棉花嫁接后通过水平基因转移获得的遗传物质,到第3代后便由于性状分离而无法稳定遗传,说明通过水平转移获得的序列在多代遗传后仍存在丢失的可能[107]。因此,用嫁接诱导水平基因转移,并实现新种质的选育,还有更多的问题需要解决。
4 问题与展望
大量研究证明,水平基因转移现象对植物进化研究和农业应用有着十分重要的意义。然而,由于技术的不足,对植物间水平基因转移现象的分析也受到了限制。这些限制主要集中在以下几个方面:首先,现阶段对植物DNA提取和组装的过程中,很难排除细菌DNA的污染和避免这种污染对分析结果产生的影响;其次,目前分析常采用的BLAST和基因树、物种树结合的方法,只在亲缘关系较远的物种中比较准确。因此,通过目前的技术手段,较难判断近期或亲缘关系较近的物种间发生的水平基因转移。除此之外,虽然人们已经发现了大规模的水平基因转移事件,但我们对其发生的条件、发生过程以及转移基因的偏好性等并没有过多的了解。相信随着测序技术和生物信息学的进一步发展,基因组组装和系统发育分析的质量必将越来越高,植物间的水平基因转移现象会得到更清晰的认知和更广泛的应用。
此外,目前对植物水平转移的基因作用的研究也不够深入。虽然研究者们已经对部分转移基因的功能进行了预测,并证明了一些外源基因能够在植物体内转录,但是要想确认这些基因是否真正具有功能,还需要更多的证据。基因功能研究是科学研究的一个重要方向,对于探究生物进化和基因的利用意义重大。随着现代生物技术的发展,对于植物间水平转移的基因功能研究也必将更加深入。而对于转移基因的功能挖掘,也会帮助我们更好地利用水平基因转移。
当然,已经有研究者对水平基因转移的应用提出了一些构想。比如,有学者认为,将植物质体工程与水平基因转移技术相结合,将有助于天然营养化合物的生物合成[108]。但是,这些应用中还存在着许多问题,例如,如何实现水平转移基因的可控性,如何将水平转移的基因固定到受体植物的基因组中并实现稳定遗传和表达等。而对这些问题的研究与攻克,也必将有助于人类对水平基因转移技术的利用。相信随着科学的发展,水平基因转移必将受到越来越多的关注,成为生物基础研究和农业改良应用的有力手段。
致谢:感谢浙江大学农业与生物技术学院园艺系夏晓剑教授在本文的写作与修改过程中提出的宝贵意见。
猜你喜欢
今日农业(2021年11期)2021-08-13
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
广西林业科学(2016年1期)2016-03-20
癌变·畸变·突变(2014年1期)2014-03-01
遗传(2014年3期)2014-02-28
世界科学(2014年8期)2014-02-28
食品科学(2013年6期)2013-03-11
世界科学(2013年6期)2013-03-11
中国烟草学报(2012年2期)2012-04-09
