
甜樱桃扩展蛋白PavEXPA2基因克隆与表达分析
2023-01-19 07:45张焱仇志浪申洛男文晓鹏
华中农业大学学报(自然科学版) 2023年1期
张焱,仇志浪,申洛男,文晓鹏
贵州大学生命科学学院/农业生物工程研究院/山地植物资源保护与保护种质创新教育部重点实验室,贵阳 550025
甜樱桃(Prunus aviumL.)属蔷薇科(Rosaceae)李属(Prunus)樱亚属植物,原产欧洲及亚洲西部,果实色泽红润,且营养价值高,深受消费者的喜爱,具有较高的经济价值[1]。
扩展蛋白(expansins,EXPA)又称为细胞壁松弛蛋白,广泛存在于植物细胞组织中,是调节细胞壁伸展和松弛的细胞壁蛋白酶[2]。它首先从黄瓜细胞壁中分离[3],后来在很多植物中陆续发现了扩展蛋白。此外,扩展蛋白能够通过其他细胞壁酶相互作用,促进细胞壁降解[4]。近年来,研究发现扩展蛋白基因参与了植物的抗逆过程。Sabirzhanova等[5]发现,在干旱条件下玉米叶片中扩展蛋白基因转录水平提高,推测扩展蛋白能够特异调节细胞壁松弛。毛竹叶片中扩展蛋白基因PeEXPA2表达水平也明显升高[6]。Cho[7]利用特异启动子使拟南芥AtEXP10基因超表达促进叶柄脱落;而抑制表达则会减少叶片脱落,并证明了叶柄脱离的原因是由于分离区扩展蛋白受到诱导发生细胞膨胀、使其产生机械压力而导致的。在花器官脱落过程中,RbEX⁃PA1的表达量上升,在延缓花瓣脱落的过程中,Rb⁃EXPA1的转录水平则降低[8]。
甜樱桃在花发育、坐果及发育期间易出现幼果异常脱落现象,但有关甜樱桃扩展蛋白基因的克隆和功能分析未见报道。基于EXPA基因在植物器官脱落中的功能,本研究以甜樱桃脱落小果果柄为材料,克隆甜樱桃扩展蛋白基因PavEXPA2,并通过在线软件对其进行生物信息学分析;利用 qRT-PCR技术分析该扩展蛋白基因在不同组织以及不同胁迫处理中的表达情况,以期为阐明其在甜樱桃生长发育中的功能提供参考。
1 材料与方法
1.1 材料及取样
以甜樱桃品种“桑提娜”为材料,分别采集茎、花芽、盛开花朵、幼叶、老叶、幼叶叶柄、老叶叶柄、2个脱落高峰期正常果柄与即将脱落果柄、2个脱落高峰期正常果实与脱落果实。以叶片为材料,使用20% PEG6000和20 mmol/L NaCl溶液分别模拟干旱和盐胁迫,采集处理0、2、4、6、8 h的材料,采集后迅速用液氮速冻,后于-80 ℃中保存备用。以上材料均设置3个生物学重复。
1.2 甜樱桃PavEXPA2基因的克隆
用植物多糖多酚RNA提取试剂盒(赛诺生物科技有限公司,中国张家口)提取甜樱桃果柄总RNA,用MutiscanGO(Thermo,美国)和琼脂糖凝胶电泳对RNA质量进行检测,并用PrimeScriptTMRT re‑agent Kit with gDNA Eraser试剂盒(TaKaRa,日本)对甜樱桃果柄总RNA进行cDNA第1链合成,使用内参基因检测cDNA的完整性并于-20 ℃保存备用。
在甜樱桃基因组数据库中搜索该基因的CDS序列,利用Primer Premier 5软件设计特异引物PavEX‑PA22-F:CACATGCTGACCTGTCCTCC、PavEX⁃PA2-R: CC GCCTAACCTCCTAAC TCTAAT,提交至上海生工生物工程有限公司合成。
以cDNA为模板,利用上述合成引物进行PCR扩增。PCR反应体系为:ddH2O 3 μL,高保真mix(TaKaRa,日本)5 μL,cDNA 1 μL,上游和下游引物各0.5 μL,共10 μL。PCR反应程序为:94 ℃预变性5 min;94 ℃变性30 s,60 ℃退火15 s,72 ℃延伸10 s,35次循环;最后72 ℃延伸7 min。用1%琼脂糖凝胶电泳检测扩增PCR产物纯度,用琼脂糖凝胶回收试剂盒对目的条带进行回收,将回收产物连接到pEASY-Blunt Cloning Kit(全式金,北京)并转化DH5α大肠杆菌,37 ℃活化1 h后吸取100~200 μL活化产物涂布于Kan抗性的LB平板上,于37 ℃过夜培养,待长出菌落后挑取单菌落进行菌落PCR验证,对检验出的阳性克隆进行培养,将阳性菌液送至上海生工生物工程有限公司测序。
1.3 PavEXPA2基因的生物信息学分析
用NCBI在 线 分 析 工 具ORF Finder(http://www.ncbi.nlm.nih. gov/gorf/gorf.Html)寻找DNA的开放阅读框,通过DNAMAN软件进行蛋白质翻译、NCBI Blastp进行蛋白质同源性比对,利用MEGA5.1构建进化树。利用在线软件TMHMM Server v.2.0(http://www. cbs.dtu.dk /services/TMHMM/)对PavEXPA2蛋白的跨膜结构域进行预测,用ProtParam(https://web.expasy.org/ prot‑param/)分析PavEXPA2蛋白的分子质量和等电点等基本理化性质,用在线分析工具SignalP 4.1 Serv‑er(http://www. cbs.dtu.dk/services /SignalP-4.1/)分析PavEXPA2基因所编码的氨基酸序列的信号肽,利用Cell PLoc2.0(http://www.csbio. sjtu.edu.cn/bioinf /Cell-PLoc-2/)进行亚细胞定位预测。
1.4 qRT-PCR分析
根据克隆获得的PavEXPA2基因序列,用prim‑er 5设计特异性引物,引物序列PavEXPA2-Y-F:CTTCTTTCTCATCTCCTCTGCC,PavEXPA2-Y-R:CCAAGGAACCATACC CACAA,在上海生工生物工程有限公司合成。以甜樱桃不同组织的cDNA为模板,以PavRSP3和PavEF1-α2为内参基因[9],引 物 序列 为:PavEF1-α2-F:ATCCAGAG‑TAGCA GAACCAATCAC,PavEF1-α2-R:GT‑TAGGCATCCAGTCCCAGAAT,在CFX 96TM Real-Time System(Bio-rad,美国)实时荧光定量PCR仪上进行。反应采用三步法,程序为:94 ℃预扩增10 min;94 ℃ 变性15 s,60 ℃退火30 s,72 ℃延伸30 s,40个循环。采用2-ΔΔCt法计算基因的相对表达量。
2 结果与分析
2.1 甜樱桃PavEXPA2克隆与生物信息学分析
以甜樱桃果柄cDNA为模板,通过PCR扩增获得与预期目的基因片段大小一致的条带,将目的条带回收并测序,结果显示序列长度为1 035 bp,开放 阅 读 框(ORF)为852 bp,编 码283个 氨 基酸(图1)。

图1 PavEXPA2基因ORF序列及编码的氨基酸序列Fig.1 ORF sequence and encoded amino acid sequence of PavEXPA2 gene
采用ProtParam分析可知,PavEXPA2蛋白质分子式为C1366H2125N3750408S15,等电点为8.90,分子质量约为30.81 ku。PavEXPA2蛋白总的负电荷残基数(Asp+Glu)为21个,正电荷残基数(Arg+Lys)为29个,由此推测该蛋白带正电荷。该蛋白含量最丰富的氨基酸分别为丙氨酸Ala(9.9%)、甘氨酸Gly(8.8%)、丝氨酸Ser(8.8%)、亮氨酸Leu(8.8%)和赖氨酸Lys(6.4%)。蛋白的不稳定系数为41.88,这表明该蛋白属于不稳定蛋白质。氨基酸残基疏水性总和(GRAVY)是-0.139,所以该蛋白亲水性较强。
采用SignalP 4.1 Server在线预测得知PavEX‑PA2蛋白存在1个信号肽,且信号肽裂解位点位于41~42(图2)。用在线软件TMHMM Server v.2.0预测跨膜结构域,结果显示,PavEXPA2为跨膜蛋白,含有2个跨膜螺旋结构(图3)。利用Cell PLoc 2.0 在线软件对甜樱桃PavEXPA2蛋白进行亚细胞定位预测分析,发现该蛋白主要位于细胞壁中。
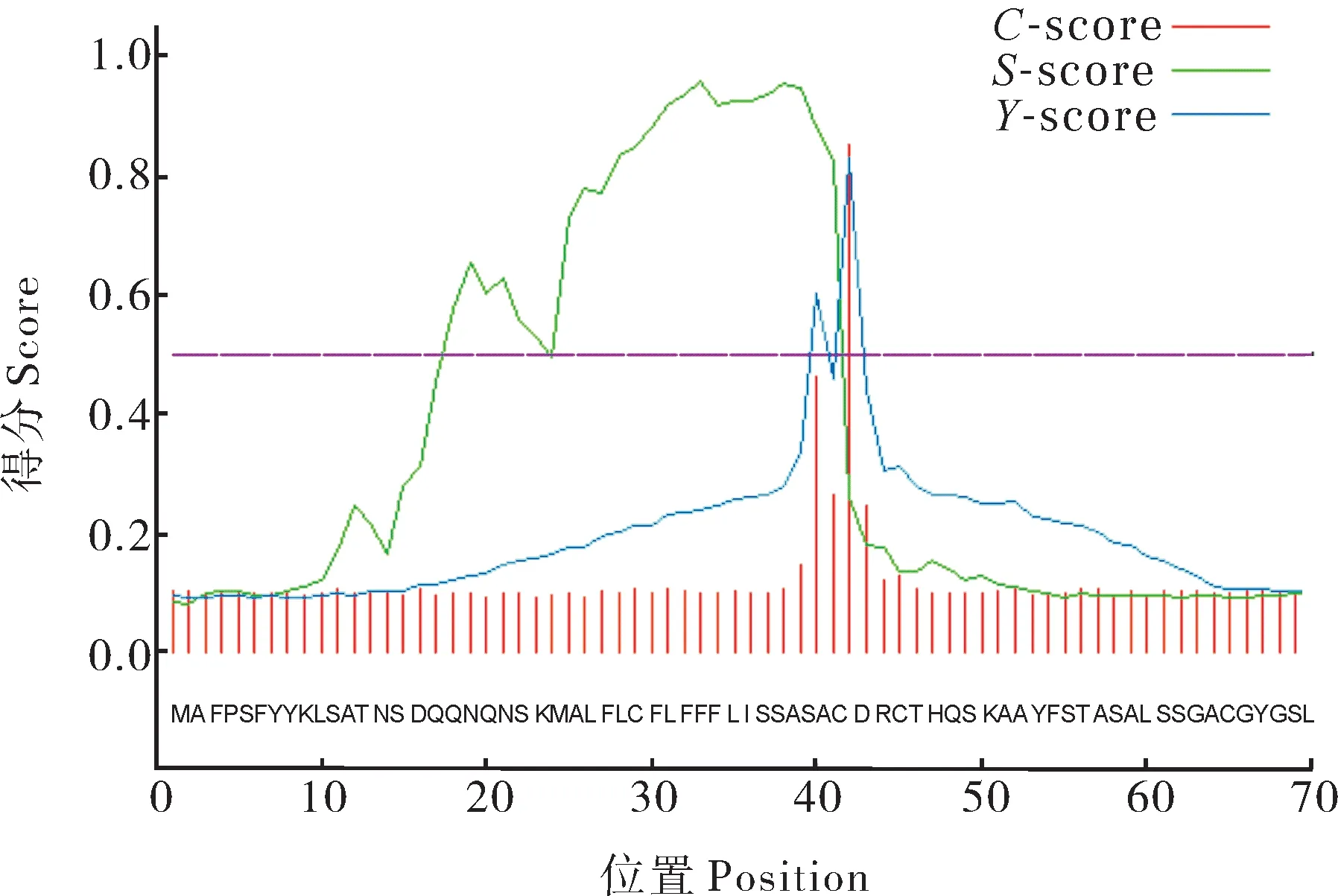
图2 PavEXPA2蛋白的信号肽预测Fig. 2 Signal peptide prediction of PavEXPA2 protein
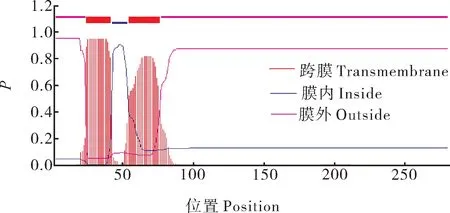
图3 PavEXPA2 蛋白的跨膜域预测Fig. 3 Prediction of the transmembrane domain of PavEXPA2 protein
2.2 PavEXPA2基因的同源性分析
将PavEXPA2基因序列在NCBI网站上进行BLAST同源检索,发现它与很多植物都具有较高的同源性,其中与扁桃(Prunus dulcis)、桃(Prunus persi⁃ca)、梅(Prunus mume)扩展蛋白基因同源性最高,分别为97.49%、97.47%和96.91%,并且发现PavEX⁃PA2有较高保守性,含有DPBB_1保守结构域。
利用DNAMAN软件将PavEXPA2基因的开放阅读框翻译成氨基酸序列,并将其序列与30个和PavEXPA2蛋白同源性较高的氨基酸序列进行对比,发现该基因编码氨基酸序列与桃、扁桃和梅扩展蛋白氨基酸序列的同源性较高,分别为95.58%、95.56%和94.76%。
利用MEGA5.1软件对甜樱桃扩展蛋白PavEX‑PA2与BLAST检索得到其他物种(柑橘、杨梅、胡杨、杨树)扩展蛋白氨基酸序列全长,构建系统进化树,结果发现甜樱桃PavEXPA2与PmEXPA2(梅)、PdEXPA2(扁桃)和PpEXPA2(桃)序列相似程度最高,亲缘关系最近(图4)。
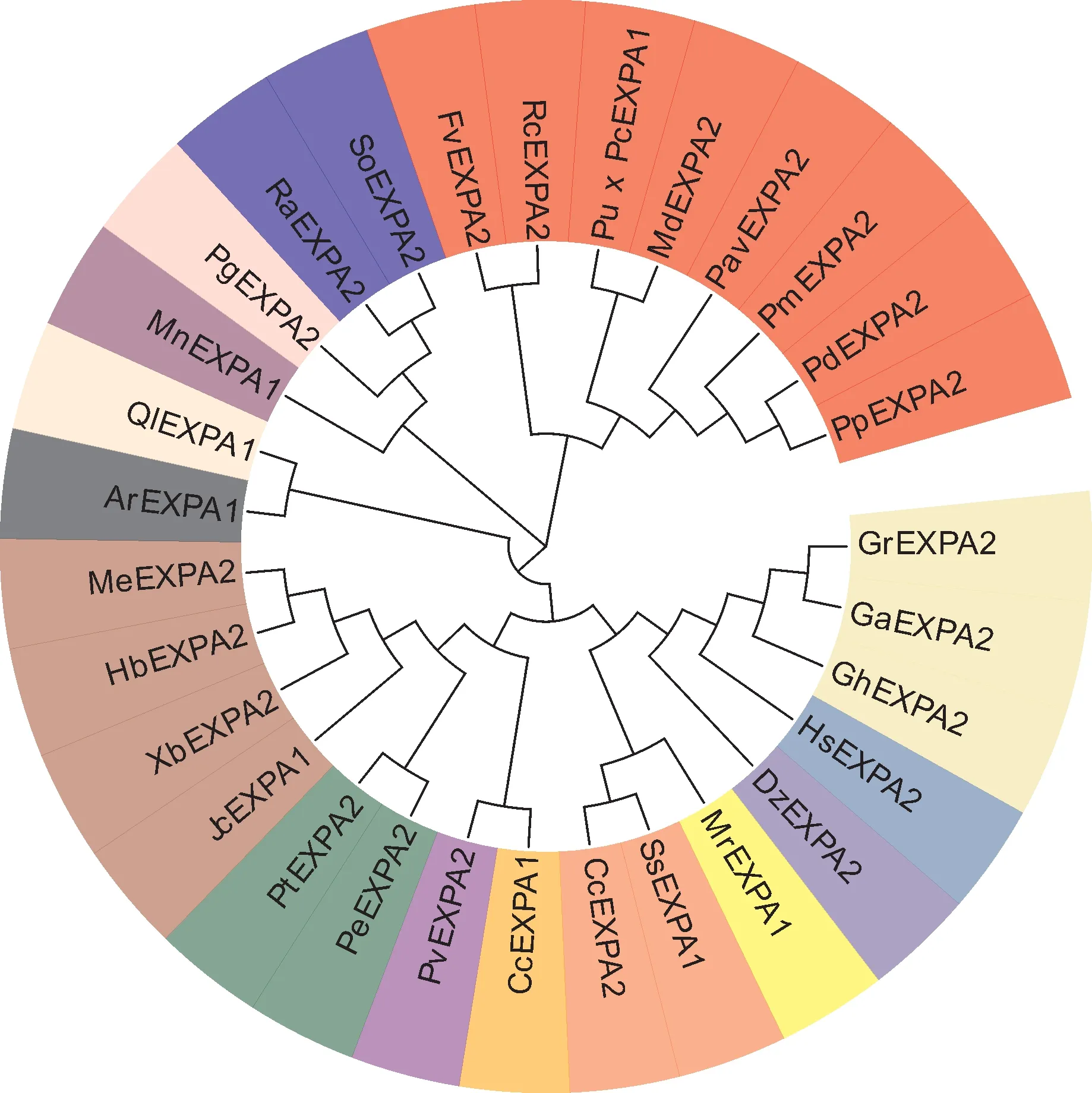
图4 甜樱桃PavEXPA2系统进化分析Fig. 4 Phylogenetic tree analysis of sweet cherry PavEXPA2 and the α-expansins of other plant species
2.3 PavEXPA2的组织特异性表达
PavEXPA2在甜樱桃“桑提娜”即将脱落组织,如茎、花芽、花朵,叶、叶柄、果柄和果实的表达结果显示,在成熟叶叶柄中表达量最高,随后依次是盛开花朵、第2落果高峰即将脱落果柄、成熟叶、第1落果高峰即将脱落果柄、第2落果高峰即将脱落果实,在第1落果高峰正常果实中表达量最低。而在第1落果高峰正常果柄、第2落果高峰正常果柄、花芽、第1落果高峰即将脱落果实、第2落果高峰正常果实、幼叶叶柄、幼叶以及茎中基本检测不到表达(图5)。因此,PavEXPA2基因主要是在成熟叶及其叶柄、脱落果柄和盛开花朵中表达,且在这些组织的表达与其成熟和脱落进程大致同步,由此推测该基因可能参与甜樱桃组织脱落的调控,尤其与叶柄脱落相关。
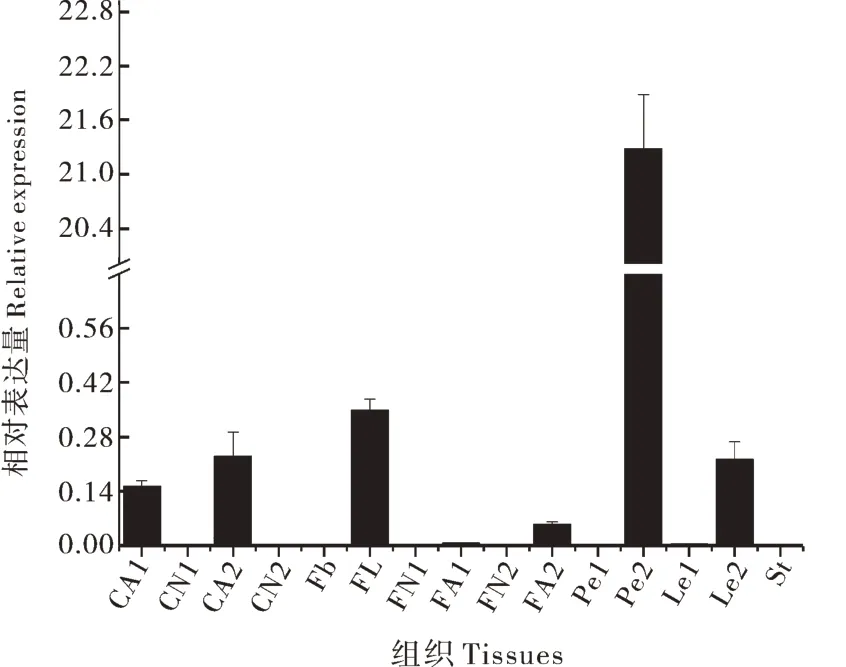
图5 甜樱桃PavEXPA2基因组织差异性表达分析Fig.5 Differential expression of PavEXPA2 gene in sweet cherry
2.4 PavEXPA2对非生物逆境的应答
在20% PEG6000和20 mmol/L NaCl处 理后PavEXPA2基因的相对表达量结果(图6)显示,在干旱处理(20% PEG6000处理)条件下,PavEXPA2基因表达量逐渐上升,在处理6 h时的表达量达到峰值,同时盐胁迫(20 mmol/L NaCl处理)下其表达量也在6 h达到峰值。而在两者表达量均达到峰值时,干旱胁迫下该基因的表达量显著高于盐胁迫,推测甜樱桃在抵御干旱及盐胁迫过程中,通过PavEX⁃PA2基因上调表达而响应逆境胁迫。表明PavEX⁃PA2在甜樱桃抵御非生物胁迫时可能起着关键作用,尤其是对于干旱胁迫的响应较为明显。
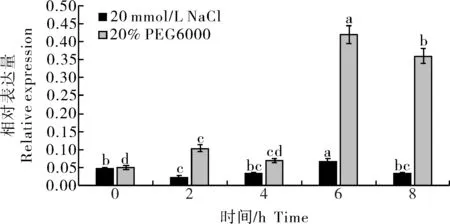
图6 盐处理和干旱胁迫下甜樱桃PavEXPA2基因的表达Fig.6 Expression of PavEXPA2 gene in sweet cherry under salt and drought treatments
3 讨论
本研究从甜樱桃克隆了1个扩展蛋白基因家族成员PavEXPA2,其编码蛋白与桃、扁桃、梅等扩展蛋白氨基酸序列有较高的同源性,PavEXPA2参与甜樱桃器官脱落的调控,尤其是叶柄脱落;在干旱及盐胁迫下,PavEXPA2基因均上调表达。因此,甜樱桃可能通过该基因上调促进脱落而抵御逆境胁迫。
扩展蛋白是1个庞大的多基因家族,目前植物扩展蛋白一般分为4类,分别为α-expansin(EXPA)、β-expansin(EXPB)、类α-expansin(EXLA)和类β-ex‑pansin(EXLB)[10-11]。扩展蛋白基因在植物基因组中广泛存在,但在基因家族序列的组成、数量、功能等方面,不同物种间有很大差异[12]。研究较多的是α-expansin和β-expansin,越来越多研究表明,在植物整个生长发育进程中扩展蛋白(尤其是α-扩展蛋白)几乎都有参与。在种子萌发、根毛起始和延长、茎和叶生长发育、叶柄脱落、花粉管延长以及果实成熟等过程中都有扩展蛋白参与[4]。另外,Downes[12]在大豆中同样发现了1个β型扩展蛋白基因CIM1,该基因通过软化柱头与花柱细胞的细胞壁、协助花粉管伸长生长,从而协助花粉管通过花柱进入子房,显示出扩展蛋白功能的多样性。
3.1 PavEXPA2基因与植物器官脱落
在其他物种中,例如在草莓[13-14]、枇杷[15]及葡萄[16]等植物中,扩展蛋白与果实的成熟软化相关。Cho[7]在拟南芥叶以及叶柄基部分析表明,扩展蛋白参与了拟南芥叶柄的脱落过程。此外,Tucker等[17]的研究也发现在大豆脱落叶柄离区中扩展蛋白有显著上调。西洋接骨木(Sambucus nigraL.)的花脱落过程中能观察到离区中扩张蛋白的多克隆抗体标记逐渐增加,并且在脱落之前的黄化阶段检测到高水平的扩展蛋白,且离区中编码扩展蛋白基因SniExp2和SniExp4上调表达[18]。因此,扩展蛋白在植物器官脱落过程中发挥了重要作用。
本研究克隆得到1个甜樱桃中的扩展蛋白Pav‑EXPA2,通过qRT-PCR发现PavEXPA2在成熟的器官及易脱落的组织中高表达,生物信息学分析结果表明,此基因可能在细胞壁中发挥作用。许多研究表明,扩展蛋白可重塑细胞壁,破坏纤维素和半纤维素之间的氢键[19-20]。细胞壁是植物细胞连接的部分,在植物器官脱落的过程中,同样涉及细胞壁的变化[21]。本研究中甜樱桃PavEXPA2在一些易脱落或者趋向脱落的器官中均有较高的表达量,说明该基因可能与甜樱桃组织的脱落相关。扩展蛋白在细胞中的功能机制已经较为明晰,但是该基因如何参与甜樱桃器官脱落尚需进一步深入研究。
3.2 PavEXPA2基因与非生物胁迫
扩展蛋白在响应非生物胁迫中有重要作用,在烟草中过表达TaEXPA2基因能提高植株对盐[22]、干旱[23]和镉[24]的抗性。同样的,在烟草中过表达TaEXPB23基因,提高了烟草的抗氧化和盐胁迫[25]的能力。在干旱条件下,小麦胚芽鞘扩展蛋白基因的表达上调[26-27]说明干旱条件下,扩展蛋白基因的功能发生了变化。Dai等[28]在拟南芥中过表达月季扩展蛋白基因RhEXPA4,增强了植株的抗旱性。在高盐条件下,玉米叶片细胞的扩展蛋白基因ZmEX⁃PA1的表达上调[29],上述研究都表明扩展蛋白与植物的抗逆性相关。本研究中PavEXPA2表达量在干旱和盐胁迫下均上调,与以上研究结果一致,说明该基因在甜樱桃抗逆过程中同样有着关键作用。
由于贵州的寡日照及喀斯特高原的特殊地理条件,樱桃的生长及果实的发育有着一定的限制。甜樱桃在幼果阶段的异常脱落已经是制约贵州甜樱桃发展的一个严峻问题,而关于甜樱桃生理落果的分子信号机制仍旧比较模糊。在逆境胁迫时, 扩展蛋白主要通过调节植物细胞壁的组分以增加细胞壁的柔韧性从而缓解胁迫对细胞造成的压力[30]。细胞的变化在逆境胁迫和脱落中存在相似之处,对扩展蛋白基因的功能研究有助于在细胞层面解析甜樱桃幼果异常脱落的机制。
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31
今日农业(2022年15期)2022-09-20
湖南农业科学(2022年5期)2022-06-22
小天使·四年级语数英综合(2022年6期)2022-06-21
今日农业(2021年11期)2021-11-27
作文·小学低年级(2021年4期)2021-11-02
作文小学中年级(2021年4期)2021-05-07
农业与技术(2021年5期)2021-03-31
河北果树(2020年4期)2020-11-26
山东农业工程学院学报(2019年3期)2019-05-11
